Arx: Civilization Upgraded. The Decentralized Blueprint for a Free Society.
True advancement does not mean erasing our differences. We reject weeding out divergent traits. an "anomaly" today may be vital tomorrow. we foster 'good diversity' to preserve the vast spectrum of human traits, just as nature intended.
Arx Revolution

genetic concepts

Human Genetics and Population Genetics
Heritability of Complex Traits
The majority of traits central to individual and group performance—general cognitive ability, impulse control, stress resilience, physical robustness, temperament, and key personality dimensions—are substantially heritable. Twin, adoption, and genome-wide association studies consistently yield heritability estimates ranging from 0.4 to 0.8. Environmental factors influence expression, but they do not negate the dominant genetic contribution. Arx regards these traits as selectable features of lineages rather than random or interchangeable variation.
Polygenic Architecture and Additive Effects
Complex traits are polygenic, arising from the cumulative, largely additive action of thousands of genetic variants, each contributing small effect sizes. Modern polygenic risk scores and genome-wide polygenic scores enable reasonable prediction of trait distributions across individuals and populations. Small systematic differences in allele frequencies between groups therefore produce meaningful shifts in trait distributions at the population level. Arx understands that selection operates on the entire polygenic architecture, not on isolated genes.
Population Structure and Allele Frequency Variation
Human populations exhibit non-random genetic structure resulting from historical patterns of migration, genetic drift, founder effects, and differential selection pressures. Allele frequencies for trait-associated variants differ systematically across ancestral groups. These differences extend beyond superficial characteristics to cognitive, behavioral, and physiological outcomes. Denial of population-level genetic variation lacks scientific support. Arx views this structure as the foundational material for deliberate lineage stewardship.
Sexual Dimorphism and Sex Chromosome Effects
Males and females differ genetically (XX vs. XY) and phenotypically in ways that extend far beyond reproductive anatomy. Sex-linked genes, hormonal influences, and sex-specific imprinting produce average differences in spatial reasoning, verbal ability, risk-taking, aggression, empathy, and life-history strategy. These are evolved adaptations, not social constructs. Arx designs cohort structures and child-rearing practices that account for—rather than suppress—these biological realities.
Genetic Load, Mutational Meltdown, and Dysgenic Trends
Modern conditions—relaxed natural selection, widespread medical interventions, and altered reproductive patterns—permit deleterious mutations to accumulate across generations. This increases genetic load and reduces mean fitness in some populations. Arx counters this through intentional selection, efficient reproductive scheduling, and closed-loop environments that reduce load and elevate mean adaptive capacity over time.
High-Fitness Lineage Amplification
High-fitness lineage amplification is the deliberate process of selecting and propagating genetic and cultural traits that confer adaptive advantages: superior cognitive ability, emotional resilience, physical robustness, and behavioral stability. In Arx, this is achieved through meritocratic governance, intentional reproductive choice, and structured child-rearing environments designed to maximize cohort outcomes. The percentage of beneficial alleles in the input population directly influences long-term results: higher initial concentrations lead to compounding effects over generations as natural and conscious selection reinforce advantageous variants while minimizing deleterious ones. Unmanaged mass immigration, by contrast, can introduce rapid allelic diversity that dilutes adaptive excellence, increases genetic load, and accelerates dysgenic trends through mismatched selection pressures.
Genetic Drift and Its Effects
Genetic drift refers to random fluctuations in allele frequencies that occur in small or isolated populations. Unlike natural selection, drift is unguided by fitness and can fix neutral or mildly deleterious variants over generations. In small groups or populations with limited gene flow, drift may reduce genetic diversity or allow harmful mutations to accumulate by chance. Arx mitigates uncontrolled drift through intentional population management, merit-based selection, and structured reproductive pairing to maintain adaptive allele frequencies.
Epigenetics and Heritable Environmental Influence
Epigenetics introduces a layer of heritable modification that does not alter the underlying DNA sequence. Environmental factors—nutrition, chronic stress, toxins, or lifestyle—can influence gene expression through mechanisms such as DNA methylation and histone modification. These epigenetic marks can persist across one or more generations, effectively transmitting environmental experience to offspring without changing the genetic code. Arx accounts for epigenetics by engineering stable, high-quality environments that promote beneficial epigenetic profiles, reinforcing adaptive gene expression in lineages.
Natural Selection and Unconscious Drift in Modern Contexts
Natural selection acts as the primary unguided filter of evolution, favoring alleles that enhance survival and reproduction in specific environments. In ancestral conditions, strong selective pressures eliminated deleterious variants and amplified adaptive ones. In modern contexts, however, relaxed selection—driven by medical advancements, reduced mortality, and altered reproductive patterns—allows deleterious mutations to accumulate. This contributes to unconscious drift: a passive erosion of mean fitness over time. Arx counters this trend by replacing passive drift with conscious, directed selection.
Conscious Evolution as Active Stewardship
Humans are the first species capable of understanding their own evolutionary history and deliberately shaping their future trajectory. Conscious evolution, as implemented in Arx, replaces unguided processes with intentional stewardship. Through merit-based pairing, cohort optimization, closed-loop environmental engineering, and cryptographic governance, Arx applies directed selection to amplify beneficial traits actively. This ensures lineages compound adaptive excellence rather than degrade through random or relaxed pressures, transforming evolution from a passive drift into a purposeful ascent.
The Self-Reinforcing Loop of Genes, Culture, and Environment
Genes interact dynamically with culture and environment in a self-reinforcing feedback loop. Genetic predispositions shape cultural norms—populations with high average impulse control may foster disciplined societies, while those with elevated creativity drive innovative traditions. These cultures, in turn, mold environments: structured, high-trust societies create selection pressures that favor compatible traits, such as cognitive acuity in knowledge-driven economies. Over generations, the environment reinforces the genes that thrive within it, perpetuating the cycle. Arx leverages this loop intentionally: by designing closed-loop habitats with meritocratic cultures, it generates pressures that enhance adaptive genetics, ensuring lineages evolve upward rather than regress.
These concepts form the scientific foundation of Arx. The project does not seek to deny, erase, or equalize variation. It seeks to recognize, measure, and amplify what has already proven viable—voluntarily, sovereignly, and without apology.
Founder Effect

The Compounding Consequences of Founder Effect
Founder Effect
The Founder Effect is a specific manifestation of genetic drift that arises when a small group of individuals—the “founders”—separates from a larger source population to establish a new one. These founders carry a limited subset of the original genetic variation. As the new population grows and multiplies, allele frequencies in the descendants are shaped disproportionately by whatever variants happened to be present in that tiny founding sample rather than by the broader diversity of the parent population. By genetic drift or conscious evolution, alleles can surge to high frequency purely by chance, or by design, while others are lost entirely. Over generations, certain alleles may reach fixation (frequency of 1.0), becoming the sole version of that gene in the entire population.
This mechanism is closely related to, but distinct from, the population bottleneck effect. Both involve a sharp reduction in effective population size that amplifies either random genetic drift or conscious evolution and reduces diversity, but the bottleneck occurs when an existing large population is suddenly decimated (by disaster, famine, or persecution) before rebounding. The founder effect, by contrast, is the establishment of an entirely new colony by a handful of migrants. In both cases, the genetic “hand” dealt (or selected) to the survivors or colonists largely determines the health, cognition, appearance, and adaptive capacity of all future descendants. Healthy founding populations compound excellence; flawed ones compound deficits.
Unmanaged gene flow into established populations produces parallel dysgenic consequences. A population with a mean IQ of 100 that experiences sustained mass immigration from groups averaging IQ 85 will see its overall cognitive distribution shift downward over generations. The same dilution occurs with physical robustness, immune function, metabolic efficiency, facial symmetry, and other polygenic adaptive traits. Arxes are deliberately engineered to stabilize against these pressures. By selecting high-fitness founders and enforcing voluntary, merit-based membership standards, Arx communities harness the founder effect as a tool of conscious evolution rather than leaving outcomes to random drift or external demographic forces.
Real human populations vividly illustrate the phenomenon. Northern European lactase persistence—the ability of adults to digest milk—spread rapidly after small founding groups in dairy-farming regions carried the allele; founder dynamics helped amplify it alongside strong selection. Conversely, Arab tribal lineages display dozens of population-specific recessive mutations (beta-thalassemia, certain glycogen storage diseases, Familial Mediterranean Fever) amplified by geographic isolation and cultural endogamy. Founders carrying deleterious variants can seed entire future populations with disease; the risk is sharply magnified by consanguinity, as closely related carriers are more likely to produce homozygous offspring. Yet moderate relatedness among founders (not extreme inbreeding) can be advantageous: it accelerates the establishment and fixation of rare beneficial alleles in the new gene pool. Beneficial founder effects also occur—e.g., the CCR5-Δ32 HIV-resistance mutation rose in frequency in Northern European-descended groups partly through medieval founder dynamics.
The founder effect is therefore a double-edged engine of rapid evolutionary change. When guided by deliberate, high-agency selection of founder DNA, it becomes a powerful accelerator of adaptive excellence. Arxes exist precisely to wield this force intentionally—choosing lineages that compound health, intelligence, resilience, and beauty rather than allowing dilution or decay. In this way, Arx transforms what has historically been an unguided process into purposeful, sovereign human ascent.
Beneficial Founder Effects and Adaptive Alleles
Beneficial founder effects follow a clear sequence: a small group carrying a rare advantageous allele separates and establishes a new population; genetic drift rapidly elevates the allele’s frequency; and local selection pressures then strongly favor it, often in concert with cultural practices that maintain isolation. The result is a population unusually enriched for traits that enhance survival and reproduction in its specific environment. Arxes deliberately replicate this pattern through conscious selection of founders, accelerating the compounding of adaptive excellence rather than awaiting rare chance events.
CCR5-Δ32 and Pathogen Resistance
The 32-base-pair deletion in the CCR5 gene (CCR5-Δ32) disables the CCR5 co-receptor that HIV-1 uses to enter cells, conferring near-complete resistance to most strains in homozygotes and slower progression in heterozygotes. The mutation likely originated once in Northeastern Europe or the Western Eurasian Steppe several thousand years ago (estimates range from ~2,500 to over 6,000 years). It rose to appreciable frequencies (up to 10–16% in Northern Europe, with a north–south cline) partly through founder effects in small medieval or earlier populations, possibly amplified by selection from historical pathogens (speculation includes smallpox, plague, or other viruses, though not HIV itself). The allele remains largely absent outside European and some West Asian populations, demonstrating how drift in isolated founder groups can elevate a protective variant that later proves valuable against new threats.
Arctic Adaptations: FADS and Related Variants in Inuit Populations
Inuit and related Arctic groups descend from small founding populations that entered harsh, high-fat, marine-mammal diets with limited plant-based precursors for long-chain polyunsaturated fatty acids (PUFAs). Strong signals of positive selection appear in the FADS gene cluster (FADS1, FADS2, FADS3) on chromosome 11 and in CPT1A. These derived alleles alter fatty-acid desaturation and metabolism, reducing the body’s conversion of short-chain to long-chain omega-3 and omega-6 PUFAs—adaptive when the diet already supplies abundant pre-formed long-chain fats from seal, whale, and fish. The variants also associate with lower height and weight in some contexts and influence lipid profiles. Nearly fixed or at very high frequency in Greenlandic Inuit due to founder effects and intense selection in small, isolated groups, these alleles exemplify metabolic fine-tuning to extreme environments.
Duffy Null Allele and Malaria Resistance
The Duffy-null allele (FYO or FYB-33, a promoter mutation disrupting DARC expression on erythrocytes) prevents Plasmodium vivax (and P. knowlesi) from using the Duffy antigen receptor for cell invasion. It is nearly fixed across much of sub-Saharan Africa (reaching >90–100% in many West and Central African populations) due to strong positive selection. While the allele may have existed at low frequency as standing variation, its dramatic rise and fixation in ancestral African populations reflect intense pressure from vivax malaria. Founder-like dynamics in expanding or isolated tribal groups, followed by selection, made this one of the most striking examples of a protective allele becoming population-defining. In modern contexts it continues to influence vivax malaria epidemiology, though some Duffy-negative individuals can still experience low-level infection.
Lactase Persistence in Northern Europe
One of the clearest examples is adult lactase persistence—the continued production of the lactase enzyme that allows digestion of lactose in milk. In most human populations, lactase expression declines after weaning. In Northern Europe, a regulatory mutation (primarily -13910*T in the MCM6 gene upstream of LCT) arose in small founding groups associated with early dairy farming roughly 7,500 years ago in the region between the central Balkans and central Europe. Founder dynamics amplified the allele, and strong positive selection followed as milk provided a reliable caloric and nutrient source during periods of famine and disease. Today the trait reaches frequencies of 80–95% in many Northern European-descended populations, illustrating how a rare variant in founders can become nearly universal.
High-Altitude Adaptations in Andean Populations
Andean highlanders, living above 3,500 m for millennia, show elevated hemoglobin concentrations as a primary response to chronic hypoxia. Variants in genes such as EGLN1 (encoding PHD2, a key oxygen sensor) and others in the HIF pathway contribute to this phenotype. Specific haplotypes (e.g., certain EGLN1 combinations) appear enriched in Andean groups, likely amplified through founder effects in early high-altitude settler populations and subsequent selection for oxygen-carrying capacity. Unlike the Tibetan pattern of blunted erythropoiesis (driven heavily by EPAS1), Andeans tolerate higher hemoglobin levels, which enhances oxygen delivery but carries trade-offs such as increased blood viscosity and higher risk of chronic mountain sickness. These population-specific genetic signatures again highlight how small founding groups encountering novel pressures can rapidly shape adaptive physiology.
Immune Variants such as HLA-B*27
Certain HLA alleles, including variants of HLA-B27, illustrate how founder effects can elevate rare immune-recognition molecules. HLA-B27 is known for both increased risk of autoimmune conditions (e.g., ankylosing spondylitis) and potential protective effects against specific viral infections through enhanced antigen presentation. Frequency differences across populations partly trace to historical founder events and drift in small groups, with subsequent selection modulating infectious disease outcomes. Small populations frequently amplify such immune variants, which may later prove advantageous or costly depending on the pathogen landscape.
In all these cases, the founder effect served as an evolutionary accelerator: limited initial genetic variation made rare beneficial alleles visible to selection far more quickly than in large, diverse populations. Arxes are designed to harness this same principle intentionally. By carefully choosing founders enriched for high-fitness, health-promoting, and adaptively valuable alleles—while maintaining standards that prevent dysgenic dilution—Arx communities aim to compound cognitive, physical, metabolic, and immune excellence across generations. This transforms what has historically been a matter of geographic or cultural accident into deliberate, sovereign evolutionary stewardship.
Maintaining Adaptive Genetic Variance in Selected Gene pools
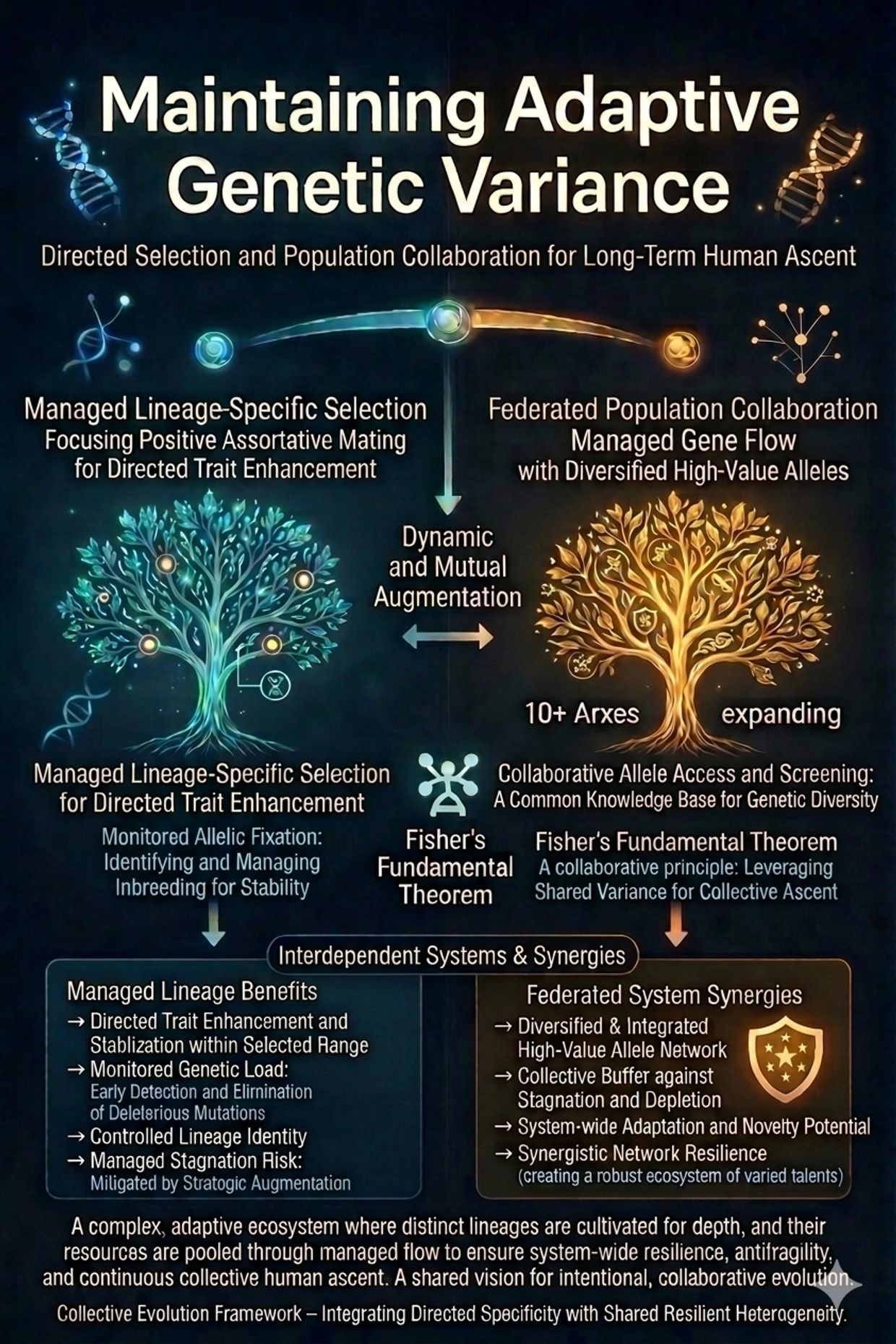
Maintaining Adaptive Genetic Variance in Large Selected Gene Pools
While strong founder effects and assortative mating powerfully amplify high-fitness traits, long-term lineage success also requires sufficient genetic variance to sustain continued adaptation. Extremely uniform gene pools, although stable in the short term, risk reduced evolvability — the capacity to respond to future environmental, pathogenic, or technological challenges. As Fisher’s Fundamental Theorem highlights, the rate of increase in fitness is proportional to the existing genetic variance in fitness. Maintaining some adaptive heterogeneity is therefore essential for antifragile, multi-generational ascent.
This does not endorse “diversity for diversity’s sake.” Random or ideologically-driven gene flow from lower-fitness source populations typically introduces net genetic load, dilutes mean trait values, and undermines the very compounding excellence Arx seeks to achieve. Unmanaged diversity is overwhelmingly dysgenic. What matters is deliberate, selected beneficial diversity: the strategic inclusion of high-value alleles, haplotypes, and polygenic combinations that enhance overall fitness without compromising core standards of health, cognition, resilience, and agency.
Arx’s federated model of multiple independent arcologies provides an ideal architecture for balancing these forces. Rather than relying on a single monolithic gene pool, numerous Arxes can experiment with different founding populations and selection criteria. Some may emphasize particular strengths — metabolic efficiency, immune robustness, cognitive specialization, or creative variance — creating a distributed portfolio of adaptive strategies. This parallel experimentation allows natural and conscious selection to identify which combinations prove most successful over decades and generations.
To prevent gradual stagnation while preserving sovereignty, Arxes can periodically and selectively incorporate screened gametes (sperm and ova) from exceptional individuals in the outside world. This controlled introgression — governed by rigorous, transparent merit-based standards — refreshes adaptive variance, introduces novel beneficial variants, and mitigates the long-term risks of overly closed populations. Far from diluting Arx lineages, such carefully managed gene flow strengthens them.
This approach creates a virtuous cycle. Successful Arxes not only compound excellence internally but also serve as net exporters of high-fitness genetic material back into the broader human population through future out-migration or gamete contribution. In this way, the Arx network simultaneously pursues internal perfection and contributes to the elevation of global human genetic potential.
By intentionally managing both the amplification of excellence and the maintenance of beneficial heterogeneity, Arxes transform genetic variance from a source of randomness into a cultivated resource — one that fuels sustained conscious evolution rather than undirected drift.
Effective Population Size (N_e)
Census population size (the total number of individuals) often differs dramatically from effective population size (N_e) — the size of an idealized population that would experience the same rate of genetic drift and inbreeding as the actual one. N_e is reduced by unequal reproductive success, overlapping generations, population fluctuations, or skewed sex ratios. A community of 1,000 residents may have an N_e closer to 100–200 if only a subset reproduces successfully or if variance in family size is high.
Low N_e accelerates genetic drift, fixation of alleles (beneficial or deleterious), and inbreeding depression, even in seemingly sizable groups. For Arx arcologies, this makes management of reproductive patterns and membership standards critical. Closed sovereignty helps, but internal dynamics — such as meritocratic incentives for high-fitness reproduction and norms that encourage broad participation in lineage continuity — are required to keep N_e robust.
By consciously maintaining adequate effective size alongside strict quality controls, Arxes avoid the pitfalls of uncontrolled small-population dynamics while still harnessing the founder effect and directed selection for rapid adaptive gains.
Fisher’s Fundamental Theorem of Natural Selection
Fisher’s Fundamental Theorem states that the rate of increase in mean fitness of a population due to natural selection equals the genetic variance in fitness present at that time. In other words, the speed of adaptive improvement is directly proportional to the amount of heritable variation available for selection to act upon.
This creates a subtle tension for intentional communities. Extreme homogenization through tight founder selection or excessive uniformity can reduce additive genetic variance, thereby slowing future evolutionary responsiveness even if current mean fitness is high. Arxes must therefore balance the stabilizing benefits of reduced deleterious variation with the retention of sufficient adaptive variance for ongoing response to novel challenges — whether environmental, pathogenic, or technological.
In practice, this means founder groups should be excellent yet not identical, and Arx governance should encourage mechanisms (such as measured assortative mating and controlled gene flow within standards) that preserve useful variation without introducing dilution. Fisher’s insight reinforces that conscious evolution is not a static optimization problem but a dynamic process: Arxes engineer initial conditions for high mean fitness while safeguarding the variance that fuels continued ascent across generations.
Mutation-Selection Balance
Mutation-selection balance describes the equilibrium reached when new deleterious mutations enter a population at a steady rate while natural selection removes them. In large, randomly mating populations with strong purifying selection, load remains relatively stable. When selection relaxes — as occurs in modern contexts through medicine, reduced mortality, and altered reproductive patterns — deleterious variants accumulate faster than they are purged, shifting the balance toward higher genetic load and eventual mutational meltdown (Muller’s ratchet in extreme cases, especially under limited recombination).
This dynamic underscores why one-time founder selection is insufficient. Without ongoing, intentional pressure favoring higher fitness, mutation inevitably erodes mean adaptive capacity over generations. Arx counters this through closed-loop habitats, merit-based reproductive norms, and structured pairing that maintain efficient purging of load while preserving beneficial variation. Conscious evolution replaces passive balance with active stewardship: continuous, measured selection that keeps deleterious input in check and allows beneficial alleles to compound.
By managing mutation-selection dynamics deliberately, Arxes prevent the slow fitness decay observed in open, low-selection societies and instead engineer sustained upward trajectories in health, resilience, and cognitive potential.
Assortative Mating (Positive vs. Negative)
Positive assortative mating occurs when individuals preferentially form partnerships with others similar to themselves on heritable traits such as cognitive ability, personality, education, height, or ancestry components. Negative assortment pairs dissimilar individuals. Humans exhibit strong positive assortment on many fitness-related traits under natural conditions.
The effect is to increase phenotypic and genetic variance in the next generation. By concentrating similar alleles, positive assortment produces more individuals at the distributional extremes — more exceptional high performers as well as more individuals at the lower tail — compared to random mating. This amplifies the impact of existing variation without immediately changing allele frequencies, serving as one of the most powerful accelerators of trait compounding in small or intentional populations.
Arx communities can harness positive assortative mating through cultural, social, and institutional design that brings high-agency, high-fitness individuals together voluntarily. Merit-based cohort formation, shared standards of excellence, and transparent evaluation systems naturally foster such pairing without coercion. When combined with careful founder selection, this mechanism dramatically accelerates the upward shift in polygenic traits central to Arx’s vision of lineage stewardship.
Careful calibration remains essential: excessive assortment on narrow traits can exacerbate pleiotropic trade-offs or reduce overall variance. Arxes therefore promote holistic fitness matching that compounds broad adaptive excellence while maintaining the variation needed for long-term resilience.
Genetic Correlations and Pleiotropy
Genetic correlations arise when the same genes or linked variants influence multiple traits, often through pleiotropy — the phenomenon in which a single gene or variant affects seemingly unrelated phenotypic outcomes. A variant that boosts cognitive performance or metabolic efficiency, for example, may simultaneously modulate immune response, stress reactivity, or disease susceptibility. Because most complex traits are polygenic, these correlations are widespread and can produce both synergistic benefits and unanticipated trade-offs.
In natural populations, pleiotropy constrains or channels evolutionary change: selection for one dimension of fitness inevitably pulls others along. For Arx communities, this demands holistic rather than narrow optimization. Selecting founders or encouraging pairings based solely on one high-value trait risks elevating correlated costs elsewhere. Conversely, when correlations are understood and leveraged, amplifying a core suite of adaptive traits (cognitive acuity, physical robustness, emotional stability) can create mutually reinforcing excellence across domains.
Arxes mitigate risks by tracking multi-trait outcomes over generations and designing environments that minimize harmful expressions of pleiotropy while magnifying beneficial ones. This layered awareness turns potential constraints into opportunities for compound fitness gains, ensuring that conscious evolution enhances overall lineage viability rather than trading one strength for hidden weaknesses.
Linkage Disequilibrium (LD) and Genetic Hitchhiking
Linkage disequilibrium refers to the non-random association of alleles at different loci on the same chromosome. When selection strongly favors one variant (a selective sweep), nearby alleles “hitchhike” along, rising or falling in frequency due to physical linkage rather than their own effects. This can preserve beneficial gene blocks or inadvertently drag deleterious variants to higher frequency until recombination breaks the associations.
In founder events or small intentional populations, LD patterns can persist longer, amplifying both opportunities and risks. Beneficial founder alleles (such as CCR5-Δ32 or lactase persistence regulatory variants) often arrive with linked haplotypes that may confer additional advantages or hidden costs. Arx protocols for founder selection and ongoing monitoring account for these haplotype blocks, using genomic tools to evaluate net fitness contributions rather than isolated variants.
Over time, managed recombination and conscious pairing allow beneficial LD to be maintained where useful while breaking apart harmful associations. This layered genomic awareness enables Arxes to steer hitchhiking effects toward compounding excellence rather than unintended load.
Norm of Reaction
The norm of reaction describes the range of phenotypes a single genotype can produce across different environments. The same genetic makeup may yield high performance in one context (nutrient-rich, low-stress, high-stimulation Arx habitat) and markedly different outcomes in another. This gene-by-environment (G×E) interaction adds essential nuance to discussions of heritability and epigenetics: “high-fitness” genotypes are not always necessarily universally superior but are often tuned to specific environments.
Arx designs closed-loop arcologies precisely to create stable, optimized environments that elicit the most adaptive expressions from selected genotypes. Engineering consistent nutrition, social trust, cognitive challenge, and minimal toxic stressors allows high-value genetic architectures to realize their full potential while minimizing harmful plastic responses.
This concept reinforces that lineage stewardship involves dual optimization: selecting superior genetic starting material and crafting the environmental conditions in which it flourishes. Arxes do not merely accumulate alleles; they curate the reaction norms that translate those alleles into realized excellence across generations.