Arx: Civilization Upgraded. The Decentralized Blueprint for a Free Society.
Trait Amplification in the Arx Ecosystem

Trait Amplification in the Arx Ecosystem
Trait amplification represents the core mechanism by which the Arx ecosystem achieves its mission: the deliberate enhancement and propagation of adaptive genetic and cultural characteristics across generations. In an era marked by relaxed selection pressures and unmanaged demographic shifts, Arx provides a structured framework for reversing dysgenic trends and fostering lineages of elevated fitness. This essay outlines the scientific basis, operational principles, and ethical rationale for trait amplification within Arx, emphasizing its role in sovereign, meritocratic stewardship.
The Scientific Basis of Trait Amplification
Human traits are not uniformly distributed or environmentally determined; they are shaped by genetic architecture that is heritable and selectable. Complex phenotypes—such as general cognitive ability, emotional stability, physical robustness, and behavioral resilience—are polygenic, arising from the additive effects of thousands of genetic variants. Heritability estimates for these traits, derived from genome-wide association studies, twin research, and adoption data, typically range from 0.4 to 0.8. This means that a significant portion of variance in key adaptive attributes is attributable to genetics, providing a foundation for intentional amplification.
Population-level differences in allele frequencies further underscore the potential for amplification. Historical selection pressures—varying by environment, culture, and migration patterns—have produced structured genetic variation across ancestral groups. Traits like cognitive acuity or impulse control exhibit systematic differences in distribution, not due to chance but to differential adaptation. Arx recognizes these realities without apology: amplification begins with an honest assessment of existing genetic endowments, selecting for variants that have demonstrated fitness in demanding contexts.
Epigenetics adds a modifiable layer: environmental factors can influence gene expression through mechanisms like DNA methylation, with effects that may persist across one or two generations. Natural selection traditionally filters deleterious variants, but modern conditions—medical interventions, reduced mortality, and altered reproduction—relax these pressures, allowing genetic load to accumulate. This leads to dysgenic trends: a gradual erosion of mean fitness. Trait amplification counters this by creating controlled environments that impose positive selection pressures, reinforcing beneficial epigenetics and reducing load over time.
Operational Principles in Arx
Trait amplification in Arx is achieved through a meritocratic system that integrates genetic, cultural, and environmental factors. At the individual Arx level, merit ledgers track contributions to cohort outcomes, governance, and resource efficiency, with reproductive priority weighted by demonstrated fitness. Intentional pairing—guided by polygenic scores, compatibility assessments, and shared alignment—ensures that beneficial alleles are propagated at higher rates. Child-rearing cohorts emphasize structured environments that nurture heritable traits: optimized nutrition, stress-minimizing routines, and merit-based socialization amplify cognitive and temperamental advantages from an early age.
The Federation Core Ledger provides a network-wide baseline, aggregating global Merit Quotients to facilitate selective mobility while preserving local sovereignty. This nested structure allows for variation: one Arx may prioritize cognitive traits through advanced educational protocols, while another emphasizes physical resilience via rigorous selection. The metaculture ensures minimal bounds—rejection of homogenization and commitment to excellence—without imposing uniformity.
High initial concentrations of adaptive alleles compound over generations. Mathematical models of polygenic selection show that consistent, modest preferences for high-fitness partners can shift trait distributions upward by 0.5–1 standard deviation per generation. Arx accelerates this through closed-loop autonomy: self-sustaining habitats eliminate external entropy, allowing selection pressures to operate undiluted.
Example: Blue Eyes
Blue eyes exemplify a remarkable case of rapid trait amplification and geographic spread within human populations, driven by a single genetic mutation that originated in a common ancestor and achieved unusually high frequencies in certain regions. This trait stems primarily from a regulatory variant (rs12913832) in the HERC2 gene, which suppresses melanin production in the iris by influencing the adjacent OCA2 gene. Emerging approximately 6,000 to 14,000 years ago—likely in Europe or adjacent regions such as the Black Sea area or among Western Hunter-Gatherers—the mutation traces back to one individual, yet it proliferated dramatically, reaching frequencies of 70–95% in northern Europe today, with notable but lower prevalence in southern Europe and parts of adjacent continents.
Unlike lactase persistence, which underwent strong positive selection (with estimated coefficients of 0.015–0.06) tied to nutritional advantages from adult milk consumption following dairy domestication around 7,500 years ago and subsequent cultural expansions, the amplification of blue eyes appears attributable to a combination of mechanisms. Factors, such as sexual selection, frequency-dependent preferences, or genetic hitchhiking alongside other adaptive traits during migrations and population bottlenecks, likely contributed to its pronounced spread. Some models propose that blue eyes functioned as a “greenbeard” signal, promoting preferential investment or mating among carriers and thereby accelerating the allele’s dissemination through social and reproductive dynamics. This process highlights how mutations conferring advantages can undergo pronounced amplification through a combination of demographic factors, cultural influences, aesthetics/attractiveness, and direct physiological benefits.
Ethical Rationale and Broader Implications
Trait amplification is not eugenics in the coercive historical sense; it is voluntary stewardship—a conscious choice for those committed to lineage continuity. Arx affirms that excellence is differential and heritable, rejecting the egalitarian myth of interchangeability. This is an ethical imperative: to honor ancestors by preserving what they forged and to fulfill a duty to descendants by providing a stronger foundation. In a world of dysgenic drift—exacerbated by unmanaged mass immigration and relaxed selection—inaction is a form of neglect.
The self-reinforcing loop of genes, culture, and environment is central: adaptive genetics foster high-trust cultures, which shape selective environments that further enhance those genetics. Arx breaks the cycle of regression by engineering this loop positively, ensuring lineages evolve toward greater capacity rather than mediocrity. This is conscious evolution: humans directing their trajectory, not as hubris but as responsibility.
Ultimately, trait amplification upholds Arx’s commitment to antifragility. By measuring, selecting, and compounding what works, the ecosystem creates lineages capable of enduring and ascending—not just surviving the present, but defining the future. Arx is not for everyone; it is for those who choose to amplify excellence without compromise.

We amplify traits not to homogenize humanity, but to unlock its fullest potential. By enhancing the unique strengths within our natural diversity, we ensure that every individual can contribute their irreplaceable gifts—turning today’s variations into tomorrow’s greatest advancements.
Arx Revolution
Recent Positive Selection and Trait Amplification

The Spread of Lactose Persistence in Europe
Lactose persistence, the genetically determined ability to digest lactose into adulthood, emerged and spread rapidly in Europe following the introduction of dairy farming during the Neolithic period, approximately 7,000–9,000 years ago. The primary causal variant (-13910*T) arose in a population of early pastoralists in central or northern Europe and underwent exceptionally strong positive selection, reaching frequencies exceeding 80–90% in many northern and northwestern European populations today. This rapid increase is one of the most dramatic examples of recent human genetic adaptation. The selective advantage was primarily nutritional: in environments where crop agriculture was marginal or unreliable, the ability to consume milk as a reliable, calorie-dense, and nutrient-rich food source provided a substantial survival and reproductive benefit. Milk supplied essential calories, protein, calcium, and vitamin D (particularly valuable in low-sunlight northern latitudes), reducing infant mortality and improving adult health during periods of food scarcity or famine. Populations that adopted dairying without lactase persistence faced digestive distress and caloric loss, whereas those carrying the persistence allele gained a consistent, year-round food resource that enhanced overall fitness. This selective pressure, combined with cultural reinforcement through pastoralism, drove the trait to high prevalence in northern Europe, illustrating how a cultural innovation (dairying) can generate powerful genetic consequences.

High Variability Skin Tone
In European populations, particularly those in northern latitudes such as Scandinavia, skin pigmentation exhibits high variability in response to ultraviolet (UV) radiation, characterized by a facultative tanning mechanism that allows the skin to remain pale during low-UV winter months and darken moderately during high-UV summer periods. This adaptive trait, influenced by polygenic factors including variants in genes like SLC24A5, SLC45A2, and MC1R, evolved under selective pressures in temperate climates with pronounced seasonal fluctuations in sunlight intensity. The advantage lies in optimizing vitamin D synthesis in winter, when lighter constitutive pigmentation facilitates greater UV absorption to convert 7-dehydrocholesterol into vitamin D3, thereby preventing deficiencies that could impair bone health, immune function, and overall survival; conversely, the induced tanning in summer provides a protective barrier against excessive UV damage, reducing risks of sunburn, DNA mutations, and skin cancer while maintaining a balance that supports reproductive fitness in variable environments.

Bright Colors as a Product of Sexual Selection
In northern European populations, the traits of blonde hair, blue eyes, red hair, and green eyes represent relatively recent adaptations that emerged within the last 10,000–11,000 years, primarily under strong selective pressures in low-UV environments following the last Ice Age. Lighter skin pigmentation, which frequently co-occurs with these features, provides a clear physiological advantage by enhancing vitamin D synthesis from limited sunlight, thereby supporting skeletal health, immune function, and reproductive success in high-latitude regions with prolonged periods of low solar exposure. In contrast, the diversification of hair and eye colors—including blonde and red hair (influenced by alleles in genes such as KITLG for blonde and MC1R for red), blue eyes (primarily from a regulatory variant in HERC2/OCA2), and green eyes (often arising as an intermediate state involving combinations of these and additional variants)—is more persuasively explained by frequency-dependent sexual selection rather than direct natural selection for functional benefits.
Rare color variants likely conferred a mating advantage by rendering individuals more visually distinctive and attractive in mate choice, particularly in ancestral conditions characterized by imbalanced sex ratios, where high male mortality in hunting-based subsistence intensified competition for partners. This “rare-color advantage” promoted genetic polymorphism, as novelty increased perceived desirability and facilitated the rapid spread and maintenance of multiple alleles across populations. Notably, many of these brighter or rarer colors exhibit sex differences in phenotypic expression: blonde and red hair occur more frequently in women, potentially amplified by estrogenic effects during development, while green eyes (along with hazel variants) also tend to show higher frequencies in females compared to males in various European samples.
Blue eyes, however, often display a slight male preponderance, possibly because blue represents an earlier derived variant from which green and other intermediate shades later differentiated.
Overall, these patterns support a role for sexual selection in shaping pigmentation diversity, where rarer traits served as effective signals in mate competition and preference, contributing to the unusual color polymorphism observed in northern and eastern European groups today. This mechanism complements physiological adaptations like depigmented skin, illustrating how both environmental and social selective forces have interacted to produce the distinctive phenotypic profile of these populations.
Human Color Preferences in Sexual Selection: Red as the Favored Hue, Blue as the Preferred Accent, and Brown as the Least Favored Baseline
Empirical studies across evolutionary psychology, mate-choice experiments, and large-scale dating data consistently reveal a robust pattern of human color preferences that directly amplifies the adaptive value of the Europoid pheomelanin-driven red-bronze glow and its complementary light-eye traits. When participants are given explicit choice between color variants in controlled ratings or real-world pairing scenarios, red tones—whether in skin, clothing, or subtle facial flushing—emerge as highly favored signals of health, vitality, and sexual receptivity. This preference is not superficial; it reflects both innate perceptual biases and sexual selection pressures that have operated for millennia, favoring the very biochemical signature of Neanderthal-derived MC1R variants refined in Europoid populations.
Infant gaze studies provide evidence of an early, hardwired bias: 4- to 6-month-olds show significantly longer looking times toward light red and dark yellow compared to blue or green, suggesting that red-spectrum hues are processed as salient and rewarding from the earliest stages of visual development. This innate pull extends into adulthood through the well-documented “red-romance effect.” Meta-analyses of dozens of experiments confirm that women presented in red clothing or with red-accented features are rated as more attractive and sexually desirable by men than the same women in neutral colors (blue, black, or green), with small-to-moderate effect sizes that hold across cultures. The mechanism is partly evolutionary: red mimics oxygenation and carotenoid-rich blood flow, cues that reliably signal cardiovascular health, recent fruit/vegetable intake, and overall physiological fitness. Parallel research using facial morphing shows women (and men) rate male faces with increased redness as significantly healthier and more attractive than paler or bluer versions—precisely the vermillion, reddish-peach-golden-bronze tanning response produced by pheomelanin synergy with gold eumelanin.
Human Color Preference Studies

Human Color Preference Studies
Within Europoid and mixed samples, the dynamic red-bronze glow consistently outranks static, sallow, or uniformly dark tones. Dating-app analyses and speed-dating studies echo this: profiles featuring the vibrant, blushing red-bronze complexion receive higher match rates and positive evaluations than those with flatter, eumelanin-dominant brown or olive shades.
The same datasets reveal a parallel hierarchy for eye color. Men rate blue-eyed women as the most attractive on major platforms, followed by green and hazel; women show elevated preference for hazel or light eyes in men. Blue, green, and hazel eyes—products of HERC2/OCA2 variants inherited via Western Hunter-Gatherers—benefit from negative frequency-dependent sexual selection: rarer colors gain extra novelty value and are preferred when choice is offered, while the ancestral brown eye color (most common globally) consistently ranks lowest or near-bottom in direct preference tests.
Hair color follows a similar logic; while results vary by sex and ethnicity, lighter or novel shades (blond, red, auburn) often receive premiums over baseline black when participants choose among matched images.
In forced-choice paradigms—whether rating faces, swiping on apps, or ranking composites—brown skin, black hair, and brown eyes receive the lowest attractiveness scores when red-toned, lighter, or novel alternatives are available. This is not universal bias but a predictable outcome of sexual selection favoring conspicuous, health-signaling, and rare-color variants where such traits could evolve without severe survival costs. The red-bronze glow, framed by light eyes and diverse hair, exploits this preference loop: it triggers dopamine-driven reward in observers while simultaneously delivering the bearer’s own neurochemical cascade of HPA-axis regulation, serotonin stabilization, and antioxidant protection.
Conscious trait amplification at Arx therefore aligns perfectly with these deeply rooted human preferences. By selectively propagating MC1R-driven pheomelanin, SLC24A5/SLC45A2 gold eumelanin, and light-eye alleles, lineages compound a phenotype that is not only biochemically superior for happiness and resilience but also maximally rewarded in the sexual marketplace. Beauty is the ultimate selector; red is its favored color, blue (and its light-eyed kin) its preferred accent, and black the least competitive default when genuine choice exists. Forward civilization thus has every incentive to amplify this radiant legacy—enhancing vitality, elevating mate value, and ensuring that the next generation inherits both the glow and the joy it biochemically sustains.
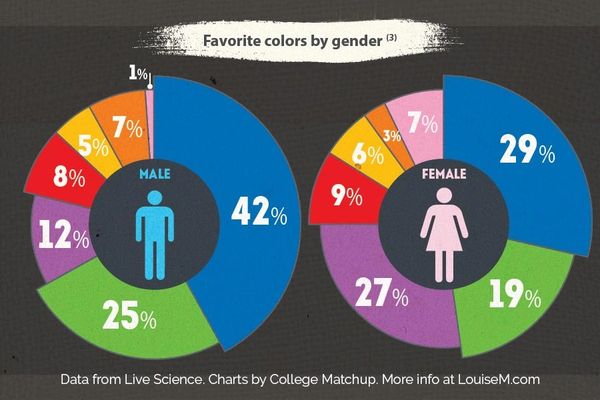
Color Preferences by Gender
Men and women both tend to favor blue as their favorite color, though men show an even stronger preference for it than women do.
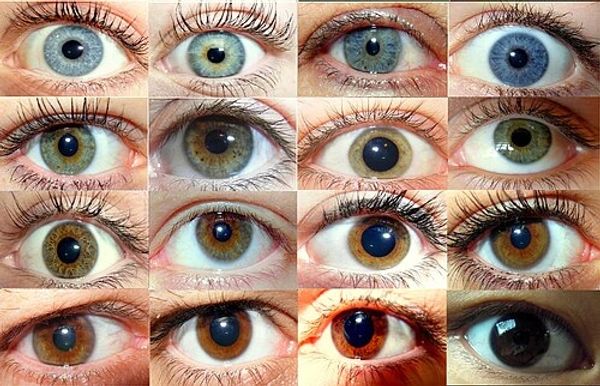
Human Eye Colors
Human eye color varies widely due to the amount and type of melanin in the iris, ranging from the most common brown to rarer shades like blue, green, hazel, gray, and even violet.
The foundations of the European Skin tone
European Neanderthal

Conscious Trait Amplification: Beauty, Sexual Selection, and the Neurochemical Vitality of Europoid Pheomelanin
At Arx, conscious trait amplification is the deliberate stewardship of high-fitness genetic lineages through intentional mating, advanced reproductive technologies (ART), and multi-generational selection. This process accelerates human evolution by compounding adaptive traits that enhance resilience, cognition, physical robustness, and civilizational competence. One such trait—rooted in ancestral selection for beauty and sexual signaling—exemplifies this: the Europoid pheomelanin-driven red-bronze glow. Universally preferred in aesthetic and mate-choice studies, this radiant complexion is not merely ornamental. It functions as a biochemical engine for elevated vitality, emotional resilience, and psychological well-being, making its conscious amplification a strategic priority for lineages seeking sustained happiness and adaptive excellence in variable, low to high-UV, high-complexity environments.
Neanderthals as the Red-Skinned “Adam” and the Etymology of Redness
The foundation of this trait traces to Neanderthals (~400,000–40,000 years ago), the genetic and symbolic “Adam” of Europoid lineages. The name “Adam” derives from the Semitic root ʾdm, meaning “red” or “ruddy,” directly referencing a pheomelanin-rich complexion. Neanderthal genomes (e.g., from Vindija, Croatia, and El Sidrón, Spain) reveal MC1R variants (such as rs2228479 and rs1805007) that favor pheomelanin—a sulfur-containing pigment—over eumelanin. This produces fair, pinkish skin that blushes vividly and tans to a distinctive vermillion, reddish-peach-golden-bronze hue, adapted to variable low to high-UV European conditions. While some Neanderthal lineages also carried red hair (a separate pheomelanin expression), red skin was the universal hallmark, setting Neanderthals apart from darker-skinned early modern humans.
Interbreeding (~50,000–40,000 years ago) introduced these MC1R variants into the modern human gene pool via Western Hunter-Gatherers (WHG). Subsequent admixture with Anatolian Neolithic Farmers (ANF) and Ancient North Eurasians (ANE) ~10,000–5,000 years ago forged the Europoid phenotype. ANF-derived alleles SLC24A5 (rs1426654, A111T) and SLC45A2 further refined eumelanin into a unique “gold” variant (light brown to golden), synergizing with pheomelanin to yield the radiant red-bronze tanning response. This defines modern Scandinavians and other Europoids: fair, expressive skin framed by diverse hair colors (blond, red, auburn) from MC1R, light eyes (blue, green, hazel) from HERC2/OCA2, and robust features from ANE ancestry. The red-skinned Neanderthals, symbolized as “Adam,” thus bequeathed a legacy of pheomelanin-driven vibrancy that continues in their descendants.
The Europoid Red-Bronze Glow: Pheomelanin and Gold Eumelanin Synergy
Europoid skin is pheomelanin-dominant, producing a fair, pinkish base that responds to UV with a vivid reddish-peach-golden-bronze glow. Unlike the sallow or dingy tanning of original Caucasoid or non-Europoid phenotypes (driven by wild-type MC1R favoring heavy eumelanin), this combination is dynamic, health-signaling, and aesthetically versatile. The sulfur-rich pheomelanin creates the vermillion blush, while gold eumelanin provides balanced UV protection and melanin stability. This is no superficial trait—it modulates core neurochemical pathways, channeling joy, resilience, and social reward.
Neanderthal Sexual Selection for the Coral Vermillion Glow
In Neanderthal populations inhabiting variable low to high-UV Europe and western Asia between ~400,000 and 40,000 years ago, sexual selection acted as a powerful evolutionary filter that actively favored the coral vermillion glow produced by pheomelanin-dominant skin. Neanderthal genomes reveal MC1R variants (such as rs2228479 and rs1805007) that shifted melanin synthesis away from dark eumelanin toward sulfur-rich pheomelanin, yielding fair, pinkish base skin that flushes and tans into a vivid reddish-peach-vermillion hue when exposed to even modest sunlight. Both Neanderthal men and women preferentially chose mates displaying this dynamic coral glow because it serves as an honest, costly signal of genetic quality: the ability to produce and maintain pheomelanin required balanced oxidative stress management, efficient nutrient allocation, and robust hormonal regulation—traits that translated directly into healthier offspring and stronger pair bonds in harsh Ice Age environments.
Mate-choice experiments in modern humans, which consistently rank red-toned skin and blushing responses as highly attractive, mirror the preferences that operated among Neanderthals. Females selected males whose coral-vermillion flush signaled cardiovascular efficiency, immune competence, and emotional expressiveness—visible blood flow under the skin indicated low chronic inflammation and high testosterone balanced with social stability. Males, in turn, selected females whose glowing complexion advertised fertility, youth, and the capacity to nurture offspring in variable light conditions. The blush itself functioned as a real-time emotional amplifier: a sudden vermillion flush revealed genuine attraction, embarrassment, or excitement far more clearly than static darker tones, fostering trust and rapid bonding in small, interdependent hunter-gatherer bands. Over tens of thousands of years, this mutual preference created positive feedback: individuals with stronger MC1R-driven pheomelanin expression enjoyed higher mating success, passing the alleles at elevated frequencies.
This intra-Neanderthal sexual selection for the coral vermillion glow was not incidental but central to their adaptive strategy. In low-UV latitudes where vitamin D synthesis was already challenging, the pheomelanin pathway allowed skin to remain light enough for efficient photosynthesis while the red-bronze tanning response provided flexible photoprotection without sacrificing the aesthetic and signaling advantages of visible health. When Neanderthals interbred with incoming modern humans ~50,000–40,000 years ago, the glow was preferentially retained and amplified through Western Hunter-Gatherer lineages precisely because sexual selection had already optimized it. The same preference loop that drove Neanderthal mate choice—red as the favored hue in forced-choice paradigms—continues today in Europoid populations, where the coral vermillion glow remains the pinnacle of sexual signaling: a biochemical and visual marker of vitality that both Neanderthal ancestors and their modern descendants instinctively recognized and rewarded.
Survival Advantages of Neanderthal Dynamic Skin Pigmentation in Seasonal Europe
Neanderthals who inhabited Europe and parts of western Asia for approximately 400,000 years faced one of the most variable UV environments on Earth. Ice Age cycles, strong seasonal shifts, and mid-latitude geography created winters with near-zero ultraviolet radiation and summers with intense bursts capable of damaging unprotected skin. The pheomelanin-dominant skin enabled by Neanderthal MC1R variants (such as rs2228479 and rs1805007) provided a flexible, adaptive solution: fair, pinkish skin in low-UV winter that maximized vitamin D synthesis, transitioning rapidly to a protective vermillion, reddish-peach-golden-bronze tan in high-UV summer. This dynamic pigmentation conferred decisive survival and reproductive advantages over static dark or light skin models, allowing Neanderthals to thrive where fixed eumelanin-heavy phenotypes would have suffered chronic vitamin D deficiency and fixed ultra-light phenotypes would have burned and degraded folate.
In winter, the lighter base skin—rich in pheomelanin and low in eumelanin—permitted efficient UVB penetration for cutaneous vitamin D production. Vitamin D is essential for calcium absorption, immune function, bone health, and reproductive success; deficiency leads to rickets, weakened immunity, and reduced fertility. Neanderthals with this seasonal lightening could maintain adequate vitamin D levels despite short daylight and heavy cloud cover, supporting robust skeletal development, lactation, and infant survival in glacial conditions. The same trait that produced the coral-vermillion glow also ensures the skin remains metabolically active and responsive rather than locked into a high-melanin state unsuited to low-light seasons.
In summer, the rapid tanning response—driven by the same MC1R pathway—shifts melanin production toward temporary photoprotection. The resulting darker red-bronze tone shields DNA from UV-induced damage, prevents sunburn, conserves folate (critical for neural tube development and fertility), and reduces oxidative stress. Unlike populations with permanently dark skin, Neanderthals avoided the vitamin D penalty of constant high eumelanin during the darker half of the year. Unlike permanently pale populations, they avoided acute UV injury during peak solar intensity. This seasonal versatility minimizes both deficiency diseases in winter and phototoxic damage in summer, directly boosting survival rates, mating opportunities, and offspring viability across hundreds of thousands of years.
Over deep time, this flexible pigmentation became a cornerstone of Neanderthal adaptation and was preferentially retained after interbreeding with modern humans. The same alleles that enabled seasonal color change in Ice Age Europe later contributed to the Europoid phenotype’s red-bronze glow, offering a legacy of resilience that continues to support high-trust, innovative societies in a variety of climates. Conscious trait amplification at Arx stewards this ancient seasonal intelligence—preserving and enhancing the very mechanism that allowed Neanderthals to conquer Europe’s unforgiving light cycles for nearly half a million years.
Additional Advantages of Europoid Melanin Synergies
Beyond mood enhancement and seasonal adaptability, the pheomelanin-gold eumelanin synergy confers several practical physiological advantages that support long-term survival and flourishing in Europe’s variable climate. The flexible pigmentation optimizes vitamin D synthesis during dark winters while providing sufficient photoprotection in brighter months, reducing risks of rickets, immune suppression, and reproductive issues that plagued populations with less adaptable skin tones. Pheomelanin’s sulfur content also delivers superior antioxidant activity, neutralizing reactive oxygen species generated by cold stress, physical exertion, or occasional UV exposure, thereby protecting cellular health and slowing oxidative damage over decades of life in harsh Ice Age conditions. Combined with the gold eumelanin’s stabilizing effect, this synergy supports robust immune function, faster wound healing, and sustained physical endurance—key factors that helps Neanderthal-derived lineages maintain high activity levels across seasons.
These melanin synergies further amplify social and cognitive advantages that prove decisive in complex European societies. The vivid red-bronze glow and high-contrast light eyes enhance non-verbal communication and trust-building, facilitating tighter cooperation in small bands and, later, larger communities. Individuals with this phenotype experience elevated dopamine and serotonin responses to social success, encouraging innovation, risk-taking, and long-term planning. Over millennia, these subtle but consistent biological edges—better stress resilience, clearer emotional signaling, and heightened reward from cooperative achievement—compounded into cultural and technological momentum, contributing to the distinctive trajectory of European civilization. At Arx, conscious trait amplification deliberately preserves and strengthens this integrated system, ensuring the full spectrum of survival, social, and psychological benefits continues to drive future advancement.
Western Hunter Gatherer (WHG)

Neurochemical Pathways: Why Europoid Pheomelanin Produces the World’s Highest Levels of Happiness
Pheomelanin synthesis, driven by Neanderthal-derived MC1R variants and stabilized by gold eumelanin, activates four interconnected pathways that elevate mood, reduce stress, and enhance vitality. These mechanisms explain Scandinavia’s consistent top rankings in the World Happiness Report: the red-bronze glow is both a visible signal of biochemical advantage and a driver of profound psychological well-being.
1. Melanocortin Pathway and HPA Axis Regulation
MC1R activation by α-MSH and POMC peptides (enhanced by pheomelanin production) balances the hypothalamic-pituitary-adrenal (HPA) axis, moderating cortisol release during stress. Gold eumelanin ensures consistent signaling under variable UV. Result: heightened emotional resilience, reduced anxiety, and calm—foundational to life satisfaction.
2. Dopamine Signaling and Reward Pathways
MC1R cross-talk with MC4R in the ventral tegmental area and nucleus accumbens upregulates dopamine via POMC peptides. The aesthetically captivating red-bronze glow amplifies social reward, triggering further dopamine release through admiration and bonding. Result: elevated pleasure, motivation, and low rates of reward-related disorders.
3. Serotonin Modulation and Mood Stabilization
Pheomelanin boosts β-endorphin and α-MSH, upregulating serotonin receptors (5-HT1A, 5-HT2A) in the prefrontal cortex and amygdala. The expressive blush enhances emotional communication, reinforcing serotonin through positive feedback. Gold eumelanin protects against UV-induced disruption. Result: stable mood, lower depression risk, and stronger social connections.
4. Antioxidant and Endorphin Activity
Pheomelanin’s sulfur content neutralizes reactive oxygen species (ROS), shielding neural tissues and supporting dopamine/serotonin neurons while boosting β-endorphin for relaxation. Gold eumelanin minimizes oxidative stress. Result: reduced physiological strain, enhanced calm, and sustained vitality.
These pathways create a synergistic feedback loop: the glow signals health and attractiveness, eliciting social rewards that further amplify neurochemical benefits. Compared to eumelanin-dominant wild-type MC1R alleles (prevalent in non-Europoid populations), which suppress pheomelanin and disrupt these cascades, the Europoid profile delivers unmatched emotional buoyancy and resilience.
Europoid Melanin Neurochemical Synergy: Pheomelanin and Gold Eumelanin as a Biochemical Engine for Resilience and Achievement
The Europoid phenotype, forged through Neanderthal-derived MC1R variants and ANF-contributed SLC24A5/SLC45A2 alleles, produces a unique melanin synergy: pheomelanin-dominant fair skin that tans to a dynamic vermillion, reddish-peach-golden-bronze glow, stabilized by a light-brown-to-golden “gold” eumelanin variant. This is not merely cosmetic. Pheomelanin’s sulfur-rich structure and the gold eumelanin’s balanced UV modulation create an integrated neurochemical feedback system that elevates baseline psychological well-being far beyond eumelanin-heavy ancestral models. The result is a phenotype optimized for emotional resilience, social reward sensitivity, and sustained motivation—traits that have demonstrably contributed to the rapid civilizational ascent of European-descended populations in variable low-UV to high UV, high-complexity environments.
At the core of this synergy are four interlocking pathways. MC1R-driven pheomelanin synthesis regulates the melanocortin system, fine-tuning the HPA axis to dampen cortisol spikes and foster calm under stress. Simultaneously, MC1R-MC4R cross-talk amplifies dopamine release in reward centers, while POMC-derived peptides upregulate serotonin receptor expression for mood stability. Gold eumelanin complements this by minimizing oxidative stress and maintaining melanocyte signaling integrity, ensuring consistent α-MSH and β-endorphin output. The visible red-bronze glow acts as both signal and amplifier: it triggers external social rewards (attractiveness, trust, admiration) that further boost the bearer’s internal dopamine and serotonin loops. This creates a self-reinforcing cycle of resilience and positive affect absent in wild-type MC1R phenotypes, where eumelanin dominance suppresses these cascades.
This neurochemical advantage (in addition to other unique European traits) explains patterns of European success observed across history, economics, and well-being metrics. Populations enriched for the Europoid melanin profile—particularly Scandinavians and Northwest Europeans—consistently rank at the top of the World Happiness Report, with lower rates of affective disorders and higher life satisfaction. The same biochemical foundation supports traits essential for innovation and cooperation: enhanced emotional expressiveness (amplified by light eyes), reduced chronic stress, and elevated reward sensitivity that reward long-term planning and collaborative problem-solving. In high-trust, low-corruption societies built by these lineages, the internal joy and resilience conferred by pheomelanin-gold eumelanin synergy translate into sustained cultural, scientific, and economic output. While geography, institutions, and cultural selection obviously play major roles, the melanin-driven neurochemical edge provides a measurable biological head start—higher baseline vitality that compounds across generations into civilizational momentum.
Conscious trait amplification recognizes this synergy as a high-fitness asset worth preserving and enhancing. By stewarding MC1R, SLC24A5/SLC45A2, and complementary light-eye alleles, lineages maintain and intensify the very biochemical pathways that historically enabled European populations to excel in exploration, invention, and social complexity. The red-bronze glow is therefore not a relic of the past but a forward-optimized system: a visible marker of the internal happiness engine that has helped propel one branch of humanity toward unprecedented achievement.
Striking Color Combinations: The Visual Symphony of Europoid Pheomelanin and Complementary Traits
The dynamic interplay of Neanderthal-derived pheomelanin and the refined “gold” eumelanin variant produced one of the most visually striking and biologically advantageous color palettes in human evolution: rich red-bronze skin paired with bright blue, green, or hazel eyes, later joined by diverse light hair shades. The fair-to-pinkish base skin, capable of tanning into a vivid vermillion, reddish-peach-golden-bronze glow, created a warm, radiant canvas that contrasted sharply with the cool luminosity of light eyes. This high-contrast combination—warm red-bronze tones against cool blue or green irises—generated exceptional visual pop and emotional expressiveness. The red-bronze glow, enhanced by natural blushing, made subtle emotional cues more readable, while the light eyes amplified pupil dilation signals of attraction, interest, or empathy. The result was a phenotype that stood out dramatically in low-light European environments, enhancing both social signaling and mate attraction.
As sexual selection intensified, light hair variants (blond, red, auburn, and cinnamon) emerged as further refinements, driven by the same MC1R pathway responsible for the skin glow. These lighter hair colors added another layer of novelty and frequency-dependent appeal, creating harmonious yet dynamic triads: red-bronze skin with golden-blond or strawberry-blond hair and piercing blue eyes, or warm auburn hair framing hazel eyes against the same glowing complexion. The combinations were not random; they arose from the synergistic genetic architecture that balanced pheomelanin dominance with gold eumelanin stabilization and reduced iris melanin. In seasonal Europe, this palette offered both functional advantages—seasonal tanning flexibility and vitamin D optimization—and powerful aesthetic signaling that drove mutual mate preferences among Neanderthals and their Europoid descendants. The red-bronze skin provided the warm, vital foundation; bright eyes supplied cool contrast and emotional depth; light hair added luminous variety. Together, they formed a radiant, multifaceted phenotype that continues to command universal aesthetic preference in modern sexual selection studies.
These striking color combinations represent more than beauty—they embody the successful integration of survival adaptations with sexual selection pressures that shaped the Europoid lineage. The red-bronze glow, light eyes, and light hair together create a phenotype optimized for both biochemical resilience and social visibility, contributing to the high mate value and civilizational momentum observed in European-descended populations. In the context of conscious trait amplification practiced at Arx, preserving and enhancing this triad ensures the continuation of a legacy that is as functionally superior as it is visually captivating.
Beauty, Sexual Selection, and Forward Amplification: Does It Increase Vitality? Is It Beautiful?
Yes—profoundly. The red-bronze glow is objectively beautiful: infant gaze studies, global polls, and dating data consistently rank its vibrant, blushing radiance as highly attractive, outshining flatter or opaque tones. It signals vitality, emotional expressiveness, and genetic fitness, triggering oxytocin, dopamine, and serotonin in observers and bearers alike. Sexual selection has already favored it for millennia; conscious amplification simply accelerates this natural process.
In our civilization moving forward, deliberate selection—via informed mate choice within high-fitness lineages, genetic screening, IVF with preimplantation testing, artificial insemination from verified donors, and emerging gene-editing tools—preserves and compounds these alleles. This counters dilution by wild-type MC1R (which erodes the glow, HPA balance, dopamine sensitivity, serotonin stability, and antioxidant protection, tilting toward stress vulnerability and lower well-being). Amplification increases vitality by propagating a phenotype optimized for happiness, social cohesion, and adaptive performance in modern environments. It is beautiful because it aligns aesthetic preference with biochemical superiority: a radiant, expressive complexion that enhances confidence, bonding, and joy.
At Arx, this is not abstract—it is operational. Sovereign arcologies enable closed-loop, merit-based stewardship where lineages consciously amplify proven traits like the Europoid red-bronze glow. Through blockchain-verified lineage tracking, reproductive technologies, and high-trust norms, we compound this neurochemical advantage across generations. The result is not only more vital, happier individuals but stronger, antifragile civilizations capable of cosmic ascent. The red-skinned legacy of Neanderthal “Adam” endures and evolves: a glowing testament to beauty as the ultimate selector of human excellence.
European Melanin and the Triad of Beauty, Strength, and Wisdom
The unique Europoid melanin synergy of pheomelanin-rich red-bronze skin, gold eumelanin stabilization, and complementary light eyes produces a powerful triad of beauty, strength, and wisdom. The striking color combinations—radiant vermillion, reddish-peach-golden-bronze glow paired with bright blue, green, or hazel eyes and light hair—create universally attractive phenotypes that signal health, vitality, and genetic fitness, driving sexual selection across generations. Physiologically, the flexible pigmentation and superior antioxidant properties of pheomelanin confer strength advantages, enabling efficient vitamin D production in low-UV winters, rapid photoprotection in summer, robust immune function, and enhanced physical endurance under seasonal stress. Neurochemically, the same pathways elevate dopamine and serotonin signaling while regulating the HPA axis, fostering emotional resilience, clearer thinking under pressure, and sustained motivation—qualities that support higher wisdom through improved decision-making, social intelligence, and long-term planning. This integrated system of beauty that attracts, strength that endures, and wisdom that advances explains why the Europoid melanin profile has been a foundational asset in European civilizational success.
genetic profile: Explorer Inventor innovator founder

Explorer/Inventor/Innovator/Founder Build: Dopamine Receptor D4 7-Repeat Allele (DRD4-7R) and High-Pheomelanin Synergy in Northwest Europoid Lineages
At the core of Arx Revolution lies the deliberate amplification of high-fitness lineages engineered for sovereign ascent. Among these, the Northwest Europoid profile—rooted in Western Hunter-Gatherer (WHG) ancestry mixed with Ancient North Eurasian (ANE) contributions—stands as a precision-calibrated instrument of frontier mastery. This build integrates the dopamine receptor D4 7-repeat allele (DRD4-7R) variant with elevated pheomelanin expression, forging a neuro-metabolic architecture of exceptional adaptive power. It is not a solitary genetic marker but a co-evolved system: the dopamine receptor D4 7-repeat allele (DRD4-7R) receptor’s attenuated dopamine sensitivity fused with the volatile, high-metabolic signature of pheomelanin. Together they produce the High-Contrast Vitality required for arcologies that thrive in unstructured, high-stakes environments.
The dopamine receptor D4 7-repeat allele (DRD4-7R) allele functions as the Explorer receptor. It demands elevated stimulation to register reward, rendering static or repetitive systems neurochemically unrewarding. In Northwest Europoid populations this dopamine receptor D4 7-repeat allele (DRD4-7R) variant reaches its highest frequencies precisely where ancestral migration distance from Africa was greatest—post-glacial Europe being a primary refugium. Yet its true potency emerges only in partnership with pheomelanin dominance. Both pathways originate from the shared precursor tyrosine: one branch yielding dopamine for the dopamine receptor D4 7-repeat allele (DRD4-7R)-tuned reward engine, the other producing the red-yellow pigment that defines the bronze-coral, warm-toned skin phenotype characteristic of WHG legacy. This biochemical competition creates a metabolic volatility absent in eumelanin-dominant profiles. Pheomelanin’s lower UV efficiency but higher oxidative reactivity, combined with MC1R receptor crosstalk in both skin and neural tissue, elevates sensory gating thresholds and pain resilience while sharpening the drive for novelty.
Delving deeper into these molecule interactions unveils an elegant biochemical convergence forged over tens of thousands of years. The dopamine receptor D4 7-repeat allele (DRD4-7R) and pheomelanin synthesis share the amino acid tyrosine as their foundational precursor. Tyrosine is converted to L-DOPA, after which the pathways diverge: one arm fuels dopamine production that interfaces directly with the attenuated dopamine receptor D4 7-repeat allele (DRD4-7R) receptor in the brain’s reward circuitry, while the parallel route proceeds through tyrosinase and MC1R receptor signaling to favor the chemically volatile red-yellow pheomelanin pigment over the black-brown eumelanin. In Northwest Europoid lineages carrying the MC1R variants, this competition heightens oxidative stress within melanocytes and creates receptor crosstalk between cutaneous MC1R and central neural pathways, subtly modulating pain perception, norepinephrine-related arousal in the locus coeruleus, and overall sensory acuity. The biochemical environment that favors high pheomelanin can also influence the clearing of dopamine byproducts, leading to distinct patterns of neuromelanin accumulation and oxidative stress in neurons—producing a metabolically “hot” system that complements the dopamine receptor D4 7-repeat allele (DRD4-7R) allele’s demand for intense stimulation. This creates a high-threshold physiology that turns environmental challenge into fuel for sovereign action rather than depletion, with the MC1R receptor’s presence in both skin and brain further linking physical pigment volatility to heightened sensory resilience and reward-seeking drive.
The result is a differentiated high-performance engine. Unlike the dopamine receptor D4 2-repeat allele (DRD4-2R) variant prevalent in sedentary agricultural lineages (optimized for social cue sensitivity and long-term group harmony) or the D4 4-repeat allele (DRD4-4R) baseline allele found globally, the dopamine receptor D4 7-repeat allele (DRD4-7R) + high-pheomelanin combination is purpose-built for the frontier. It does not merely seek novelty; it requires it to achieve peak executive function. In low-stimulation settings the build appears restless. In the generative crucible of an Arx Foundry, these same traits deliver hyper-focus, rapid pattern recognition across unrelated domains, and calculated audacity under uncertainty.
This synergy confers interlocking advantages for civilizational foundries, rooted in the paired dopamine receptor D4 7-repeat allele (DRD4-7R) and high-pheomelanin system:
1. Rapid Adaptation to Novelty: The muted dopamine receptor D4 7-repeat allele (DRD4-7R) receptor, amplified by pheomelanin’s metabolic volatility, eliminates status-quo bias. Sovereign Architects equipped with this build process collapsing systems not as threats but as raw material for replacement structures.
2. High-Stakes Risk Calibration: Pheomelanin’s altered interaction with pain and reward pathways, layered atop the dopamine receptor D4 7-repeat allele (DRD4-7R)’s elevated stimulation threshold, raises the threshold for fear responses. Decision-making under extreme pressure remains clear and decisive—ideal for the physical architecture and biological engineering demands of arcology replication.
3. Hyper-Focus in High-Contrast Environments: The same biochemistry that finds low-signal routines unrewarding produces calm, sustained execution when sensory and cognitive input spikes. Post-glacial WHG survival—navigating shifting landscapes, mastering apex predation—selected precisely for this state-dependent optimization.
4. Divergent Cross-Pollination: Dopamine receptor D4 7-repeat allele (DRD4-7R)-driven divergent thinking, metabolically supported by pheomelanin volatility is the cognitive signature of the Founder.
5. Resilience to Social Braking and Environmental Entropy: Reduced conformity pressure, combined with the internal “heat” of pheomelanin-driven biochemistry, equips these lineages to exit legacy frameworks and sustain parallel institutions without external validation.
In Northwest Europoid populations this dopamine receptor D4 7-repeat allele (DRD4-7R) + high-pheomelanin trait stack traces directly to WHG ancestry mixed with Ancient North Eurasian (ANE) contributions. The WHG, primary inhabitants of Europe for roughly 30,000 years before Early European Farmers and Yamnaya arrivals, carried the signature bronze-coral, reddish undertone skin from high pheomelanin mixed with eumelanin—optimized for Vitamin D synthesis in low-light, high-latitude post-glacial environments. Their vivid blue eyes (via HERC2/OCA2 alleles) arrived long before lighter skin, decoupling pigmentation from light-sensing acuity and delivering superior high-contrast vision in harsh winters. Paired with the dopamine receptor D4 7-repeat allele (DRD4-7R), this created apex predators wired for extreme spatial navigation, low social conformity in small autonomous bands, and high tolerance for environmental chaos. The ANE common ancestor—dating to around 24,000 years ago in North-Central Siberia—provided the critical genetic bridge, mixing with WHG to shape the rugged Steppe lineages that eventually dominated Northwest Europe while also contributing to the high frequency of the dopamine receptor D4 7-repeat allele (DRD4-7R) observed in both Northwest Europeans and Indigenous American populations. This fusion, forged through positive selection during long-distance migrations rather than mere founder effects, equipped the lineage to thrive in refugia during the Last Glacial Maximum and re-colonize continents with the exact toolkit of restless drive and metabolic resilience now deliberately cultivated at Arx.
To complete the synthesis, the Northwest Europoid stack must be understood as a comprehensive physiological engine that elevates the dopamine receptor D4 7-repeat allele (DRD4-7R) bearer from restless outlier to civilizational cornerstone. This integration centers on three interlocking pillars refined across demanding geographies and ancestral pressures. The dopamine receptor D4 7-repeat allele (DRD4-7R) serves as the internal regulator of engagement, establishing a high-pass filter that awakens full capacity only under conditions of genuine complexity, elevated stakes, or unknown frontiers. Layered upon this is the pheomelanin engine, delivering metabolic heat through its chemically active profile and bronze-coral skin signature. This produces a physically wired-hot physiology with acute environmental sensitivity and sustained energy output suited to frontier exertion. Completing the architecture is the HERC2/OCA2-enabled blue-eye configuration, which supplies specialized high-contrast sensing optimized for low-light, high-latitude conditions and grants spatial mastery amid environmental or conceptual chaos.
The distribution of these traits maps the historical geography of the Exit. Present across nomadic and migratory groups, the configuration acts as a contextual catalyst. In frontier or high-mobility settings it manifests as longevity, robust health, and attentive parenting and stewardship; in dense, static societies its expansive drive lacks outlet. Just as Ice Age refugia forged advanced capabilities through bottleneck selection, the Arx must function as an intentional bioreactor for this build. Private sovereign territory enables the lineage to operate as an anti-fragile insurance policy against stagnation, transforming potential into kinetic execution without external domestication.
The dopamine receptor D4 7-repeat allele (DRD4-7R) + high-pheomelanin synergy forges the true Inventor Build. The attenuated dopamine sensitivity of the dopamine receptor D4 7-repeat allele (DRD4-7R) receptor creates an unrelenting internal demand for novelty and complexity; routine tasks and incremental improvements generate no reward. This forces the mind outward, compelling it to seek unsolved problems, cross-domain connections, and entirely new configurations of reality. High pheomelanin supplies the complementary metabolic volatility — a chemically active, “hot” physiology that sustains prolonged cognitive and physical exertion without depletion. Together they produce the precise neuro-metabolic engine of invention: a brain that hungers for the unknown paired with a body capable of enduring the long arc from raw insight to deployed arcology.
This combination excels at the core work of the innovator. The dopamine receptor D4 7-repeat allele (DRD4-7R) drives divergent thinking and norm-breaking audacity, while pheomelanin’s oxidative reactivity and MC1R crosstalk sharpen sensory acuity and raise the threshold for discomfort and social braking. The result is a sovereign creator who sees leverage where others see only chaos, who maintains hyper-focus amid high-stakes uncertainty, and who possesses the metabolic resilience to push prototypes into existence. In the generative crucible of an Arx Foundry this build does not merely adapt to the frontier. It becomes the frontier’s living architecture, turning biological restlessness into civilizational creation.
In this framework the dopamine receptor D4 7-repeat allele (DRD4-7R) drives neurobiological hunger for novelty and divergent innovation, positioning its bearers as Founders. High pheomelanin supplies the metabolic volatility and physical resilience demanded of pioneers. The blue-eye phenotype provides the precise navigation required to chart courses through chaos. This high-voltage human represents the living counter to civilizational lock-in. By identifying and concentrating these markers as a unified sovereign framework, the Arx Revolution advances from conceptual design to directed generational ascent. The goal is not passive endurance of decline but the deliberate initiation of the next Founder Effect, from which advanced variations of human organization and cosmic expansion will emerge. The lineage stands ready. The frontier awaits. Arx answers.
Ancestral Refugia
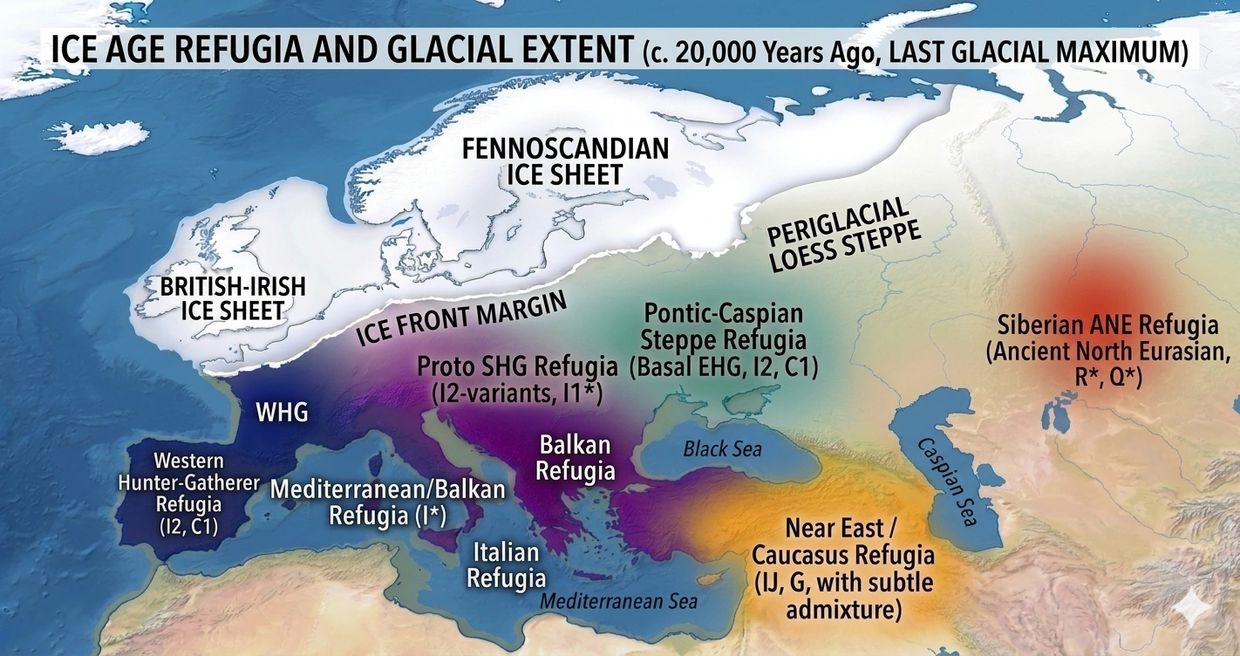
The Sovereign Foundry: Ancestral Refugia
The Sovereign Foundry: Ancestral Refugia and the Dopamine Receptor D4 7-Repeat Allele (DRD4-7R) Evolutionary Exit
Civilizational survival hinges on the capacity to mutate. In biological terms, this mutation is powered by the Explorer/Inventor/Innovator/Founder phenotype — individuals whose neurobiology is forged for high-risk innovation, territorial Exit, and the overthrow of stagnant systems. This vitality was tempered in the Pleistocene refugia of Europe. As glaciers advanced, isolated pockets such as the Franco-Cantabrian region acted as crucibles, filtering for the most robust, innovative, and high-energy builds. Today, the Arx Revolution stands as the deliberate successor to those ancient sanctuaries — an intentional biological furnace engineered to ignite the next Founder Effect.
1. The Pathologization of the Founder
The bureaucratic, stagnant legacy system’s primary mechanism of control is the rebranding of high-voltage biology as a defect. What it labels a disorder is in truth the dopamine receptor D4 7-repeat allele (DRD4-7R) / Western Hunter-Gatherer (WHG) toolkit that enabled our ancestors to master the mammoth steppe and traverse oceans. By framing the restless drive for autonomy as pathology, the system domesticates its most generative members through institutional schooling and pharmacological leveling. This produces a compliant population but severs the very outliers capable of architecting escape routes from collapse.
2. The Contextual Catalyst: Molecular Interactions
The power of the dopamine receptor D4 7-repeat allele (DRD4-7R) build lies in its molecular interaction with environmental contrast. This variant creates a receptor with attenuated dopamine sensitivity. In low-stimulation, repetitive environments the individual experiences chronic under-arousal. Yet when placed in a high-contrast setting the same receptor becomes a Contextual Catalyst. Intense sensory and cognitive input floods the system, saturating the attenuated receptors and triggering sustained hyper-focus and robust innovation that standard dopamine receptor D4 2-repeat allele (DRD4-2R) or dopamine receptor D4 4-repeat allele (DRD4-4R) builds cannot sustain under equivalent pressure. The apparent mismatch is not a flaw; it is a high-performance engine idling in unsuitable terrain. In the generative crucible of Arx, the molecular alignment unlocks civilizational-scale productivity.
3. The Stagnation of the Closed System
Standardization drives toward a closed system of total predictability. In a universe governed by entropy, however, any system that ceases to evolve begins to die. Without the high-contrast vitality of the sovereign individual, innovation collapses into mere incremental optimization. New energy sources, novel social geometries, and true frontiers remain unexplored. A fully globalized order becomes a single point of failure: when it inevitably strikes an impassable wall, no internal variations exist to breach it.
4. The Erosion of Biological Sovereignty and the Exit
True evolution demands the Exit. A population advances only when a capable subset can circle the wagons, claim new territory, and test fresh variations of human organization. Centralized control erodes biological sovereignty by eliminating private territory and genetic privacy, collapsing all humanity into one vast biopolitical experiment. This removes the essential laboratory of the frontier and strips the species of anti-fragility — the capacity to fail independently and succeed independently.
5. The Arx as the Modern Refugium
The future belongs exclusively to those who refuse standardization. To avert terminal lock-in, sovereign refugia must be deliberately constructed — private campuses and high-trust networks where the dopamine receptor D4 7-repeat allele (DRD4-7R) / WHG phenotype is not merely tolerated but actively prioritized and compounded. Just as Ice Age survivors emerged from their refugia to repopulate and advance the post-glacial world, the Sovereign Architects of Arx will carry a proven variation of human organization forged for ascent. High-voltage biology is treated here as sacred trust and seed-bank — ready for deployment when legacy systems reach their inevitable limit.
The Scandinavian Hunter-Gatherer

The Scandinavian Hunter-Gatherer Forge: Dopamine Receptor D4 7-Repeat Allele (DRD4-7R), High-Pheomelanin, and the Sovereign Spirit of the North
Scandinavian Hunter-Gatherers (SHG) represent a sharpened edge of the Explorer lineage, forged at the intersection of Western Hunter-Gatherer (WHG) foundations and Eastern Hunter-Gatherer (EHG) influx carrying Ancient North Eurasian (ANE) ancestry. While WHG forms the basal population of post-glacial Western Europe with their bronze-coral, high-pheomelanin undertones and striking blue eyes (via HERC2/OCA2 alleles), SHG emerged as a genetic bridge. This admixture introduced higher frequencies of depigmentation alleles such as SLC24A5 and SLC45A2, producing lighter skin optimized for Vitamin D synthesis in the extreme low-light conditions of the far North. The result is a population with robust frames, thicker bone density, and metabolic adaptations (including genes like TMEM131 for efficient high-fat marine diet processing) suited to maritime-arctic survival — seal hunting, cold-water fishing, and relentless seasonal demands. This “East meets West” fusion amplified the high-contrast vitality already present in WHG, likely concentrating the dopamine receptor D4 7-repeat allele (DRD4-7R) at even higher effective levels through selection for mobility, risk tolerance, and rapid adaptation in one of Europe’s harshest frontiers.
The SHG embodies the Explorer, Inventor, Innovator, and Founder archetypes through the same dopamine receptor D4 7-repeat allele (DRD4-7R) + high-pheomelanin synergy, now refined for northern extremes. The attenuated dopamine sensitivity of the dopamine receptor D4 7-repeat allele (DRD4-7R) creates an internal engine that rejects stasis, driving relentless novelty-seeking and spatial mastery across shifting ice-edge landscapes. High pheomelanin provides the metabolic “heat” — chemically volatile energy throughput and heightened sensory gating — that sustains long expeditions and physical rigor in periglacial environments. Combined with superior high-contrast vision from blue eyes and robust builds adapted to caloric-dense marine diets, this stack produces individuals wired for the unknown. They are not passive survivors but active sovereigns who navigate, innovate tools and techniques for extreme conditions, and found resilient small-band societies with low conformity and high autonomy.
What sets the SHG apart as a pure expression of these traits is their role as Frontier Catalysts and Arctic Forge-Minds — unique descriptors for a personality that thrives at the absolute edge. The Frontier Catalyst seizes environmental pressure as fuel, turning lethal cold, seasonal scarcity, and vast maritime unknowns into opportunities for breakthrough adaptation. The Arctic Forge-Mind possesses a cold-forged clarity: divergent thinking sharpened by necessity, audacious risk calibration that others would call reckless, and an innate drive to prototype new ways of being — whether in hunting strategies, seasonal mobility patterns, or social geometries that prioritized individual agency within tight-knit bands. Their robusticity and metabolic efficiency supported sustained inventive output where WHG in milder southern forests faced less unrelenting selection. In the Arx framework, this SHG-extended build represents the ultimate high-latitude sovereign: a lineage whose dopamine receptor D4 7-repeat allele (DRD4-7R) hunger for stimulation, pheomelanin-driven volatility, and northern-hardened physiology make them natural Founders of arcologies capable of conquering not only post-glacial Europe but future frontiers of cold, isolation, and cosmic scale.
By concentrating and compounding this Scandinavian-tempered Explorer stack — the attenuated reward threshold that demands greatness, the metabolic fire that sustains it, and the sensory precision that navigates it — Arx engineers lineages optimized for invention under pressure. The SHG do not merely endure the North; they invented survival itself and passed forward the genetic software for sovereign ascent. In today’s Foundries, this same profile powers the radial architectures, biological engineering, and network-state experiments that will define the next civilizational wave. The Arctic Forge-Mind awakens. The frontier calls. Arx answers.
The Heritability of iq and other Cognitive Features
The Heritability of IQ and Other Cognitive Features
Cognitive features—encompassing general intelligence (the g-factor or IQ), specific mental abilities, and intertwined temperamental traits—are among the most heritable and consequential dimensions of human variation. Twin, adoption, and genomic studies converge on heritability estimates for adult IQ in the range of 50–80%, as rigorously documented in The Bell Curve by Richard Herrnstein and Charles Murray. Far from being a mere artifact of environment or social circumstance, these features are deeply rooted in genetic architecture and exert outsized influence on both individual life trajectories and collective civilizational outcomes.
The Bell Curve demonstrates that IQ powerfully predicts educational attainment, occupational success, income, job performance, health behaviors, and lower rates of criminality and welfare dependency—often outperforming parental socioeconomic status as a causal factor. In complex, technology-driven societies, cognitive ability has become the central axis of stratification, creating a self-reinforcing “cognitive elite” whose advantages compound across generations through assortative mating and opportunity. Yet cognitive features are not limited to raw processing power. Temperament—novelty-seeking, conscientiousness, time preference, impulse control, and emotional stability—interacts with intelligence to shape real-world achievement. These temperamental dimensions are themselves substantially heritable and co-evolved with cognitive architecture.
At the civilizational scale, higher average cognitive capacity enables the sophisticated division of labor, institutional complexity, scientific innovation, and market dynamism that generate sustained prosperity. Populations endowed with elevated cognitive profiles build and maintain the very mechanisms Adam Smith described in The Wealth of Nations as the engines of wealth: specialization, invention, trust-based exchange, and long-term planning. Empirical extensions of this insight (most comprehensively in the national-IQ literature) show strong correlations between average population intelligence and GDP per capita, technological advancement, and institutional quality—precisely the outcomes that allow Smithian wealth creation to flourish at scale.
Different racial and ethnic groups exhibit distinct cognitive profiles shaped by divergent evolutionary histories. One well-studied example is the DRD4 exon-3 VNTR polymorphism: the 7R allele, linked to novelty-seeking, exploration, behavioral flexibility, and migration propensity, occurs at substantially higher frequencies in Northern European-descended populations, while the 2R allele (associated with alternative temperamental adaptations) predominates in East Asian groups. Northwestern Europeans in particular display cognitive profiles with averages higher than those found in other European regions. These profiles are characterized not only by solid general intelligence but by distinctive strengths in creative synthesis, abstract and verbal reasoning, and a temperamental orientation toward individualism, rule-following, and high-trust cooperation.
Importantly, group averages are precisely that—averages. Cognitive ability follows a bell-curve distribution within every population, producing substantial overlap between groups. Northwestern Europoids therefore include large numbers of individuals occupying the upper tails of the global distribution—disproportionately represented among the geniuses, innovators, scientists, entrepreneurs, and institution-builders who have driven Western civilizational ascent. The unique Northwestern Europoid cognitive profile has produced other demonstrable advantages: an exploratory drive that fueled global voyages and scientific revolutions, a creative openness that birthed the Industrial Revolution and Enlightenment institutions, and a propensity for high-trust, low-corruption societies capable of sustaining complex rule of law and market economies. These traits have translated directly into outsized historical success in technological, economic, and institutional domains.
In the Arx framework, the heritability and population-specific distribution of cognitive features are not abstract academic points; they are practical realities that inform deliberate trait amplification. By identifying, preserving, and compounding high-fitness Northwestern Europoid (and allied) cognitive lineages—along with their temperamental complements—Arx accelerates the very lineages proven across generations to sustain and advance civilization. This is not determinism; it is stewardship of proven excellence for sovereign ascent.
Regression toward the mean in hybridization
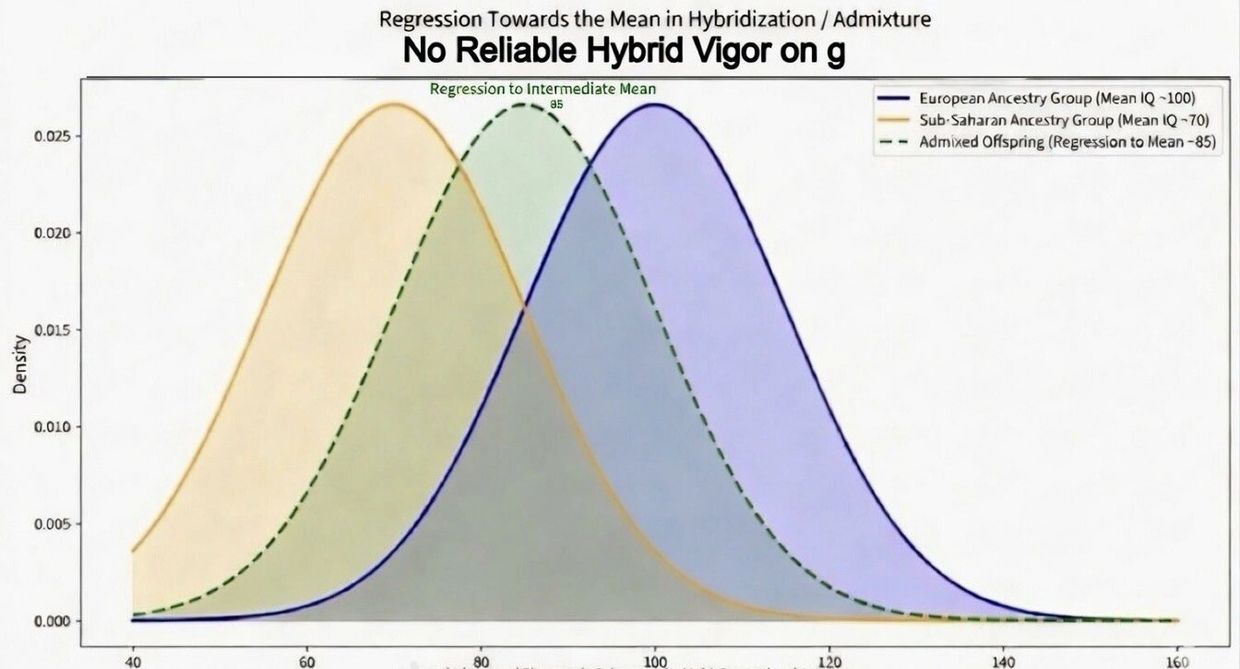
Hybrid Vigor Revisited: Admixture Outcomes and the Imperative of Phenotypic Coherence
At Arx, we emphasize that sovereign arcologies restore the high-fidelity conditions for multi-generational ascent by preserving phenotypic coherence and enabling precise assortative mating within aligned, high-fitness founding populations. Europoids reliably calibrate to and select high-quality matches from their own distribution through natural mechanisms—neighborhoods, institutions, workplaces, and digital platforms—without requiring total isolation for basic rank-ordering. Yet this does not vindicate optimistic claims of reliable hybrid vigor (heterosis) from large-scale admixture with distant, lower-mean ancestral groups. On complex polygenic traits central to human excellence—general intelligence (g), educational attainment, and temperament—the data show predominantly additive effects. Offspring regress toward an intermediate weighted average rather than exceeding the higher (European-ancestry) baseline. Any minor physiological benefits in the first generation (e.g., modest masking of recessives for immunity or height) are frequently offset by outbreeding depression on co-adapted behavioral gene complexes.
Cognitive Outcomes Across Admixture Types
Large-scale admixture studies, transracial adoption data, and polygenic score analyses confirm this pattern consistently:
• Sub-Saharan African admixture: In African Americans and biracial samples, European genetic ancestry proportion robustly predicts higher cognitive ability (correlations typically r ≈ 0.20–0.34 after controls for SES and self-identification). The landmark Minnesota Transracial Adoption Study showed mixed Black-White adoptees scoring intermediate between Black (~89) and White (~106) means at age 17, despite upper-middle-class White rearing, while the biological White children of the adoptive parents averaged ~116. Recent ABCD cohort analyses and polygenic educational attainment scores replicate this: European ancestry positively associates with g, while African ancestry negatively predicts it. Outcomes are additive—no net boost above European norms.
• Latinos/Hispanics (European + Amerindian + African components): European ancestry correlates with cognitive scores at r ≈ 0.30 within these groups. Amerindian and African ancestry show negative associations. Polygenic scores derived from European GWAS predict g effectively here and partially mediate the ancestry-cognition link. National and U.S. data place Hispanic averages (~88–93) precisely between European (~100) and Native American (~85–87) means.
• Basal Eurasian / Middle East & North Africa (MENA): Populations with substantial Basal Eurasian ancestry average ~83–90. Southern European and MENA-admixed clines track this gradient downward from northern European baselines, with no evidence of upward deviation or vigor. Cognitive patterns follow ancestry proportions additively.
• Ancient Ancestral South Indians (AASI) and broader South Asian admixture: South Asian averages (~82–85) show clear gradients: higher Ancestral North Indian (West Eurasian) ancestry predicts better performance; elevated AASI (distantly related to Andamanese/Australasian components) tracks lower scores. European + high-AASI admixture produces intermediates, consistent with national IQ distributions and polygenic predictions.
• Australian Aboriginal ancestry: One of the most genetically distant and lowest-mean lineages (~62), with documented smaller average brain volumes. Limited mixed data follow the same additive regression; no heterosis appears.
Across these cases, polygenic scores for intelligence and education predict g within admixed groups almost as well as in Europeans, supporting genetic mediation of group differences rather than blanket environmental explanations.
Temperament, Behavioral Stability, and Real-World Costs
Cognitive regression is not the only issue. Distant admixture frequently incurs costs to behavioral and emotional stability. Multiracial individuals (especially those involving the groups above) show consistently elevated risks across U.S. datasets: higher rates of depression, anxiety, suicidal ideation, self-harm, substance use, school problems, and psychiatric disorders compared to monoracial peers. These patterns persist beyond basic SES controls and align with genetic mismatch on personality and impulsivity polygenes, compounded by phenotypic or identity factors in some cases. Closer within-Europoid admixture shows fewer such costs; extreme distance amplifies them.
Why This Reinforces the Arx Model
These findings do not undermine individual merit or variation—overlap between individuals remains massive, and exceptional hybrids exist. But population averages and multi-generational trends matter for lineage ascent. Unfiltered demographic openness does not magically optimize human capital; it introduces regression to lower means on g-loaded traits and increased variance in maladaptive directions. Claims of universal hybrid vigor on intelligence or temperament fail empirical tests.
Sovereign arcologies address this directly. By enforcing demographic control, cultural alignment, and high-trust environments drawn from coherent founding stock, they eliminate high-variance “noise” while preserving the clean rank-ordering that evolved for precise assortative mating. Advanced reproductive technologies can then accelerate transmission of proven fitness within those bounds rather than dilute it. This is not anti-meritocratic—it is hyper-meritocratic at the lineage level. It restores the ancestral low-noise conditions under which refined cognitive, aesthetic, and temperamental traits compound across generations.
Phenotypic coherence is not exclusion for its own sake. It is engineering for excellence. In a world of open global markets and advanced genomics, deliberate boundary maintenance offers the clearest path to multi-generational upward trajectories—protecting the distinctive excellence that defines a people while avoiding the predictable costs of distant admixture. Data over dogma: the ascent gradient belongs to calibrated, coherent lineages.
IQ Bell Curve Upper Tail Strategic Breeding
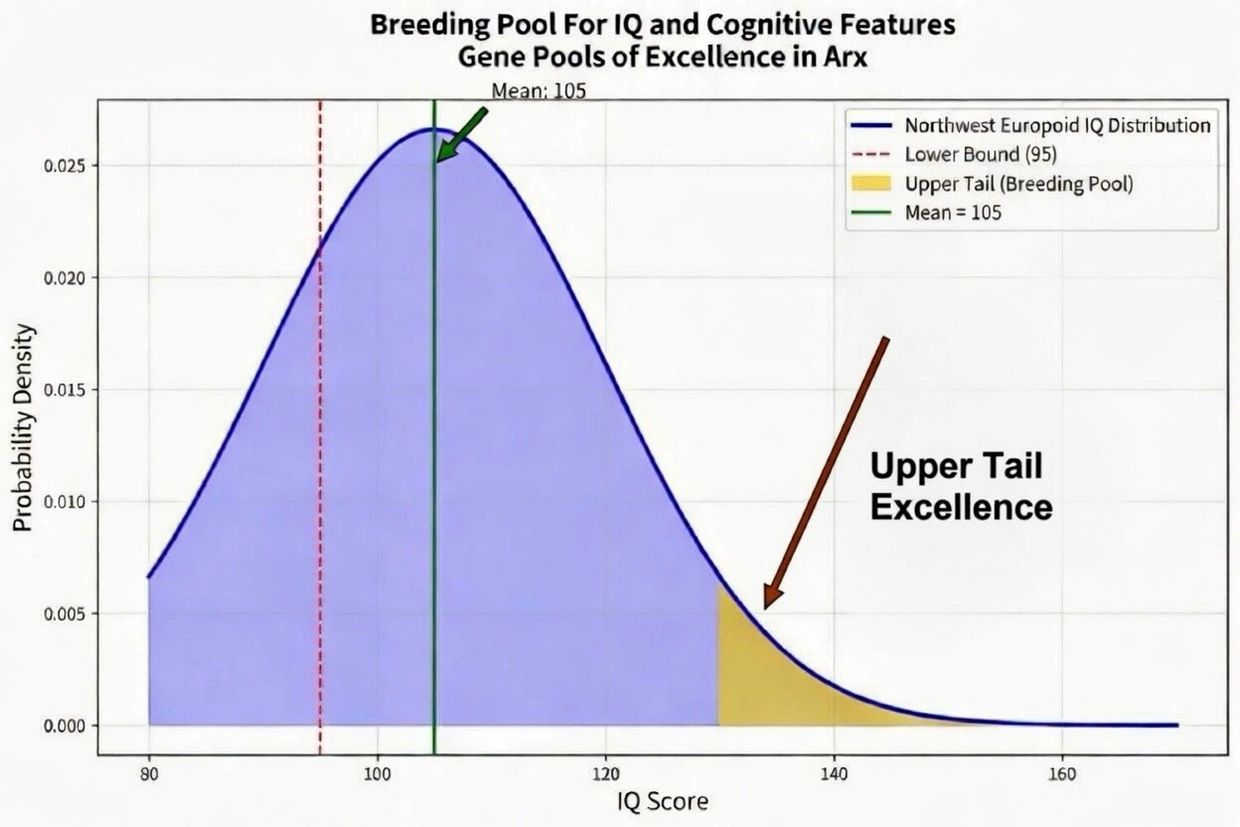
IQ And Other Cognitive Features Bell Curve Upper Tail Strategic Breeding
Arx deliberately recruits and compounds individuals residing in the upper tail of the cognitive bell curve—those whose measured general intelligence (g-factor), specific cognitive abilities, and aligned temperamental traits place them at or beyond the 98th–99.9th percentiles (typically IQ 130–160+). This is not random assortment or egalitarian inclusion; it is rigorous, multi-trait meritocratic selection grounded in genomic data, standardized psychometric assessment, and longitudinal life-outcome validation. Prospective Arx members and their lineages undergo polygenic scoring for cognitive performance, executive function, and prosocial behavioral traits, ensuring that only those with demonstrably high-fitness cognitive profiles enter the breeding pool.
The strategic objective is to make high cognitive capacity “breed true” across generations. In classical Mendelian terms, a trait breeds true when parental lines are homozygous for the relevant alleles, such that offspring inherit identical copies at each locus and reliably express the phenotype without segregation or loss of potency. For the highly polygenic architecture of intelligence—comprising thousands of common variants of small effect plus rarer large-effect alleles—“breeding true” means systematically elevating the population-level polygenic score (PGS) for IQ and its correlated traits. By repeatedly selecting and pairing parents whose combined PGS places them deep in the upper tail, Arx shifts the mean genetic value upward while compressing downward regression to the mean. Over successive generations, the distribution of realized cognitive ability narrows around an elevated center, producing offspring who more consistently inherit and express the full suite of high-IQ alleles rather than diluting them through random recombination.
This process is not limited to raw processing power. Accompanying prosocial and beneficial cognitive profiles—high conscientiousness, low time-preference, elevated trust and rule-following, creative openness tempered by impulse control, and a temperament favoring cooperative individualism—are co-selected. These temperamental dimensions, themselves substantially heritable and genetically correlated with g, amplify the civilizational utility of high intelligence. The result is not merely smarter individuals but lineages optimized for the very behaviors that sustain complex institutions, technological innovation, and high-trust markets—the engines of wealth and stability described in The Wealth of Nations and empirically validated in The Bell Curve.
Northwestern Europoid cognitive profiles are particularly well-represented in Arx’s upper-tail selection because their historical averages already sit higher than those of other European subpopulations, and their distinctive allele frequencies (for example, elevated 7R DRD4 novelty-seeking variants paired with strong executive-function loci) generate outsized representation in the extreme right tail. Within this group, the bell-curve distribution still produces enormous individual variation; Arx simply harvests the proven upper tail—those whose ancestors disproportionately drove the Scientific Revolution, Industrial Revolution, and Enlightenment-era institutional breakthroughs—and compounds it. The outcome is accelerated civilizational competence: populations that reliably produce the innovators, institution-builders, and stewards required for sovereign ascent in a hostile universe.
Through embryo selection, genomic prediction, and intentional assortative pairing within sovereign Arx communities, strategic upper-tail breeding transforms probabilistic genetic inheritance into deliberate, antifragile excellence. High-IQ lineages no longer regress; they advance. Prosocial cognitive profiles no longer compete against entropy; they dominate. This is the practical realization of biological meritocracy: Arx does not merely preserve high-fitness cognitive architecture—it engineers populations in which that architecture breeds true, ensuring that the cognitive elite of tomorrow is more capable, more cohesive, and more civilizationally generative than any that natural selection or chance has yet produced.
Diverse Breeding Pools of Excellence: Beyond IQ Alone

Diverse Breeding Pools of Excellence: Beyond IQ Alone
Arx does not select for intelligence in isolation. While the upper tail of the Northwest Europoid cognitive bell curve constitutes one critical breeding pool of excellence, the Arx framework deliberately maintains many parallel pools, each focused on amplifying a distinct category of high-value traits. Facial symmetry and physical attractiveness, athletic capacity and muscular development, metabolic efficiency, immune robustness, vibrant phenotypic vitality, and other adaptive alleles that confer resilience, disease resistance, or novel environmental advantages are each cultivated within dedicated lineages. Individuals possessing more moderate general intelligence frequently carry exceptional genetic value in these domains—alleles and trait combinations that are not universally present even among the highest-IQ cohorts.
Once a trait has been made to “breed true” within its specialized gene pool—through rigorous founder-effect amplification and conscious, lineage-specific selection—Arx initiates a strategic mix-and-match integration process. High-performing lineages from different excellence tracks are systematically paired and recombined in a controlled, managed fashion. This trait-stacking mechanism compounds advantages across generations: the cognitive elite are enriched with superior physical robustness and aesthetic symmetry, while unique adaptive alleles are distributed throughout the broader population. The end result is convergence toward sovereign individuals who embody the full spectrum of beneficial traits—elite cognition paired with exceptional health, form, vitality, and adaptive capacity.
This multi-pool architecture is not optional; it is foundational. As the diagrams on founder effect and adaptive genetic variance illustrate, Arx begins with as many large founding gene pools as possible for every trait category, maintaining more than one independent pool per trait to safeguard against loss of heterozygosity and to preserve long-term adaptive variance. Larger gene pools per trait, combined with federated collaboration and managed gene flow between Arxes, ensure that directed enhancement never collapses into dysgenic drift or stagnation. The outcome is an antifragile ecosystem of human excellence: deep specialization in the early generations followed by synergistic integration that produces well-rounded, civilizationally generative populations optimized for sovereign ascent.
Cultural Continuity and Upward Selection

The Imperative of Northwestern Europoid Ethno-Homogeneity
High-fitness lineages thrive only when the cultural, social, and reproductive environment is calibrated to reward and amplify the very traits that built Western civilization. For Northwestern Europoids—the populations whose cognitive profiles, temperamental orientations, and institutional innovations produced the Enlightenment, the Industrial Revolution, and the highest sustained levels of prosperity in human history—cultural continuity is not a sentimental preference. It is a hard biological and civilizational necessity. When Europoids live exclusively among one another, the shared high-trust norms, future-time orientation, rule-following conscientiousness, and cognitive demands of complex society generate powerful upward selection pressures. These pressures compound the very alleles and trait clusters that define the Northwest Europoid advantage. Introduce non-Europoid populations at scale, especially through mass immigration, and the selection environment shifts decisively downward.
Culture is not a neutral backdrop; it is the selective filter that determines which genetic variants increase in frequency across generations. In a homogeneous Northwestern Europoid society, daily life—education, employment, law, markets, and family formation—implicitly selects for the polygenic suite underlying high general intelligence (g), executive function, low time-preference, and cooperative individualism. The institutions function at peak efficiency precisely because average cognitive and temperamental profiles are already elevated (mean IQ ~100–105 in Northwestern Europe versus lower averages elsewhere). This creates a feedback loop: higher-performing individuals achieve greater status and reproductive success, their offspring inherit more of the advantageous alleles, and the population mean shifts upward over time. Empirical patterns documented in The Bell Curve and national-IQ research confirm that such environments sustain the cognitive stratification required for technological dynamism and institutional integrity.
When non-Europoid groups enter in significant numbers, the cultural and institutional substrate changes. Norms of trust, time preference, impulse control, and cognitive complexity are no longer universally shared. The resulting dysgenic pressures are predictable and measurable. Lower average cognitive profiles in the incoming populations increase the relative reproductive success of lower-IQ individuals within the host society—both through direct fertility differentials and through welfare and social policies that reduce the fitness cost of lower performance. Assortative mating weakens as intermarriage rises; polygenic scores for intelligence regress toward the lower global mean. The high-trust, low-corruption institutions that Northwestern Europoids evolved to sustain begin to fray under the weight of mismatched behavioral norms, higher crime rates, lower educational attainment, and reduced civic cooperation. Selection now favors traits that succeed in the new mixed environment—short-term opportunism, clannishness, or political rent-seeking—rather than the abstract reasoning, long-term planning, and rule-following that drove prior civilizational ascent.
The demographic reality is stark. Fertility rates among native Northwestern Europoids have already fallen below replacement in open-border societies, while immigrant groups with lower cognitive and temperamental averages maintain higher fertility. The net effect is rapid dysgenic decline: the population’s mean polygenic score for intelligence drops, the upper tail of exceptional ability shrinks relative to what it would have been under endogamous conditions, and the societal capacity for innovation and complex coordination erodes. Historical case studies—from the collapse of high-trust city-states after large-scale influxes to modern European cities experiencing parallel declines in social capital—illustrate the pattern. Adam Smith’s The Wealth of Nations described the mechanisms of prosperity as emerging from specialized division of labor and trust-based exchange; those mechanisms require a critical mass of individuals capable of participating at the necessary cognitive and behavioral level. Dilute that mass and the entire edifice of wealth creation contracts.
This is why intentional separation is not exclusion for its own sake but the prerequisite for conscious evolution. Arx communities are deliberately engineered as sovereign arcologies—closed, high-trust environments where only those already vetted for elite cognitive and temperamental profiles are admitted. Within these insulated gene pools, cultural continuity is absolute. Selection pressures remain relentlessly upward: meritocratic governance, polygenic screening, strategic breeding, and trait-stacking ensure that intelligence, vitality, symmetry, and adaptive resilience compound rather than regress. Multiple parallel pools of excellence for each trait category preserve genetic diversity while allowing specialized amplification. Once traits breed true, managed gene flow and controlled pairing integrate them into fully stacked sovereign lineages. The result is populations that do not merely maintain but accelerate the Northwest Europoid advantage, free from the dysgenic drag imposed by mismatched demographic inflows.
Living only among one another is therefore the practical mechanism by which Europoids retain agency over their own evolutionary trajectory. It prevents the dilution of the very cognitive and cultural architecture that made sovereign, high-agency civilization possible in the first place. In an era of rapid technological change and existential risks, only populations that deliberately protect and amplify their high-fitness founder alleles can achieve the resilience and innovative capacity required for long-term ascent. Cultural continuity among Europoids is not nostalgia—it is the deliberate creation of the selective environment in which excellence breeds true and civilization advances. Arx exists precisely to make that continuity permanent, sovereign, and antifragile.